






У казахов два аристократических рода — кожа и торе, не входящие в систему жузов. Род кожа, по генеалогическим легендам, ведет своё происхождение от Пророка Мухаммеда, его многочисленных сподвижников, четырёх первых халифов или арабских завоевателей. Торе считаются потомками Чингисхана. По данным популяционной генетики (гаплогруппы Y-хромосомы и их субклады) выполнена этническая идентификация их предков. Предки- арабы с Аравийского полуострова у кожа не выявлены. Предками большинства были «таджики». Эта популяция в прошлом занимала территории Таджикистана, Северного Афганистана, Восточного Ирана. Проживала в городах Сырдарьинского региона, возможно, Ферганской долины, а также междуречья Сырдарьи и Амударьи. Второй компонент рода кожа «монголы». Всего носителей монгольских гаплогрупп (C2, O и D) у них 6,5…31,3 %, в том числе гаплогруппы C2 — 6,5…18,8 %. Эти же два этнических компонента по частотам резко доминируют у торы. Частоты монгольских гаплогрупп 42,9…63,0 %, гаплогруппы C2 — 39,1…63,0 %. Среди носителей её субкладов не имеется потомков монголов халха. Минимально представлены потомки калмыков. Большинство является потомками калмаков и носителей этнически не идентифицированного субклада «Авары». Единственное значимое отличие торе от кожа — у них выше частоты монгольских гаплогрупп, примерно, как у казахов Старшего жуза. Данные популяционной генетики по торе и кожа не соответствуют их генеалогическим легендам.
Ключевые слова: казахи, кожа, торе, популяционная генетика, гаплогруппы Y-хромосомы, субклады, этногенез.
ВВЕДЕНИЕ
У казахов имеются два аристократических рода — кожа и торе, не входящие в систему жузов. Кожа — это казахская форма слова ходжа, в персидском языке — «господин, хозяин» (Гафуров, 1987). По генеалогическим легендам кожа ведут своё происхождение от Пророка Мухаммеда, его многочисленных сподвижников, четырёх первых халифов или арабских завоевателей. Торе считаются потомками Чингисхана. Эти представления не подтверждены данными популяционной генетики. У кожа не обнаружены носители гаплогруппы Y-хромосомы J1, являющейся мажорной у аравийских арабов, а казахи рода торе являются потомками нескольких не родственных мужчин. Целью выполненного исследования являлась этническая идентификация предков этих двух родов.
МАТЕРИАЛЫ
Инструментами популяционной генетики изучаются генетические портреты популяций, включающие гаплогруппы Y-хромосомы и соответствующие им гаплотипы (передаются по мужской линии), гаплогруппы мтДНК (по женской линии), а также аутосомные маркеры (по обеим линиям). По мужской линии торе и кожа являются обособленными популяциями. В статье рассмотрены гаплогруппы Y-хромосомы и их субклады.
Тестирование казахских торе на гаплогруппы Y-хромосомы ведётся с 2008 г. Авторы публикации (Сабитов, 2012) обобщили результаты (N=39, N — количество тестированных) по потомкам казахского хана Джанибека (1428–1480 гг.), отстоящего на 6–7 поколений от Джучи, старшего сына Чингисхана. Для шести их линий приведено число носителей мажорных гаплогрупп С3(×C3c), R2a и R1a1a. Всего 33 (6 гаплогрупп попали в «Остальные»). Их описание выполнено на основе аксиомы: «Среди казахских ханов не было документально зафиксированных фактов самозванства или усыновления. Важно отметить, что, как правило, казахские чингизиды знали друг друга […] единственным механизмом инкорпорации в состав чингизидов другой гаплогруппы мог быть адюльтер» (Сабитов, 2012: 122).
Данные из Казахского ДНК-проекта рассмотрены в публикации (Сабитов, 2014). Включают торе (N=27), кожа (N=62) и представителей 10 казахских родов (N=386). Род кожа связан «с проповедниками ислама среди казахов, которые, по всей видимости, имели различное происхождение» (Сабитов, 2014: 137). Данные по торе не проком ментированы. Как мы уже отмечали ранее, торе не являются потомками Чингисхана,
но этот вывод находится в рамках его общепринятого происхождения. В публикации (Жабагин, 2014) приведены результаты исследования, целями которого являлись верификация исторических гипотез о происхождении казахских родов торе и кожа, а также выявление степени связи между биологическим и со циальным родством их членов. Тестированы торе (N=23), кожа (N=71), представители казахских родов жалайыр (N=100) и керей (N=77), а также монгольского рода борджигин (N=88). По историческим свидетельствам, киреи и жалайыры восходят к древнемонгольским родам, которые были в тесной исторической связи с чингизидами. А Чингисхан происходит из рода борджигин. Выборка авторов публикации является научной. По торе и кожа она не пересекается с данными из Казахского ДНК-проекта. Как она соотносится с выборкой торе из публикации (Сабитов, 2012) непонятно. По результатам исследований сделаны следующие предположения:
– мажорные гаплогруппы кожа R1a, J2 и R2a указывают на регионы происхождения миссионеров — Передняя Азия, Иран и Таджикистан;
– торе и кожа имеют две общие мажорные гаплогруппы — R1a и R2a, что может быть связано с их смешением в прошлом;
– генофонды родов торе и борджигин перекрываются «по мажорной гаплогруппе C3*-M217(×M48), носителем которой, как считается, был их родоначальник Темучин (Чингисхан)» (Сабитов, 2012: 100).
В диссертации казахстанского генетика М.К. Жабагина (2017) приведены частоты гаплогрупп Y-хромосомы и их субкладов по 14 казахским родам (N=1982), включая торе (N=28) и кожа-сунак (N=107). Представляется, что данные по аристократическим родам частично опубликованы в статье (Жабагин, 2014), а новые данные — в (Zhabagin, 2017).
Данные по популяциям исторического региона Трансоксианы (бассейн Сырдарьи от Ферганской долины до её низовьев и низовья Амударьи) приведены в публикации (Zhabagin, 2017). Тестированы четыре (регионы Арыси, Шымкента, Жанакургана и Казалы) группы казахов из Туркестанской и Кызылординской областей Казахстана (N=349), а также популяции из Узбекистана: узбеки Хорезмской (N=98) и Ферганской (N=67) областей, Ташкента (N=52), дунгане Ташкентской области (N=31), каракалпаки (N=100) и туркмены (N=83) Каракалпакстана. Казахи включают торе (N=6) и кожа-сунак (N=94).
Согласно генеалогической традиции, клан кожа-сунак произошёл от мусульманских миссионеров с Ближнего Востока, принадлежавших к племени курайшитов и приходившихся родственниками по отцовской линии пророку Мухаммеду. Однако, носители гаплогруппы J1-M267, считающиеся маркером арабской экспансии, у них не обнаружены. Главный вывод авторов публикации (Zhabagin, 2017): кожа-сунак по отцовской линии происходят от нескольких неродственных местных предков. Авторы публикации предположили, что основой формирования клана явилась духовная традиция суфийских орденов Яссавия, Накшбандия и Бекташи.
Результаты исследования всех казахов Казахстана (N=1164) приведены в публикации (Khussainova, 2022). Они структурированы по родам и жузам (Старший, Средний и Младший). Отдельно приведены данные по торе и кожа. Дано только формальное описание фактических данных.
Общий объём опубликованных данных по торе и кожа приведён в таблице 1.
Таблица 1. Опубликованные данные по родам торе и кожа, а также по казахам других родов Казахстана
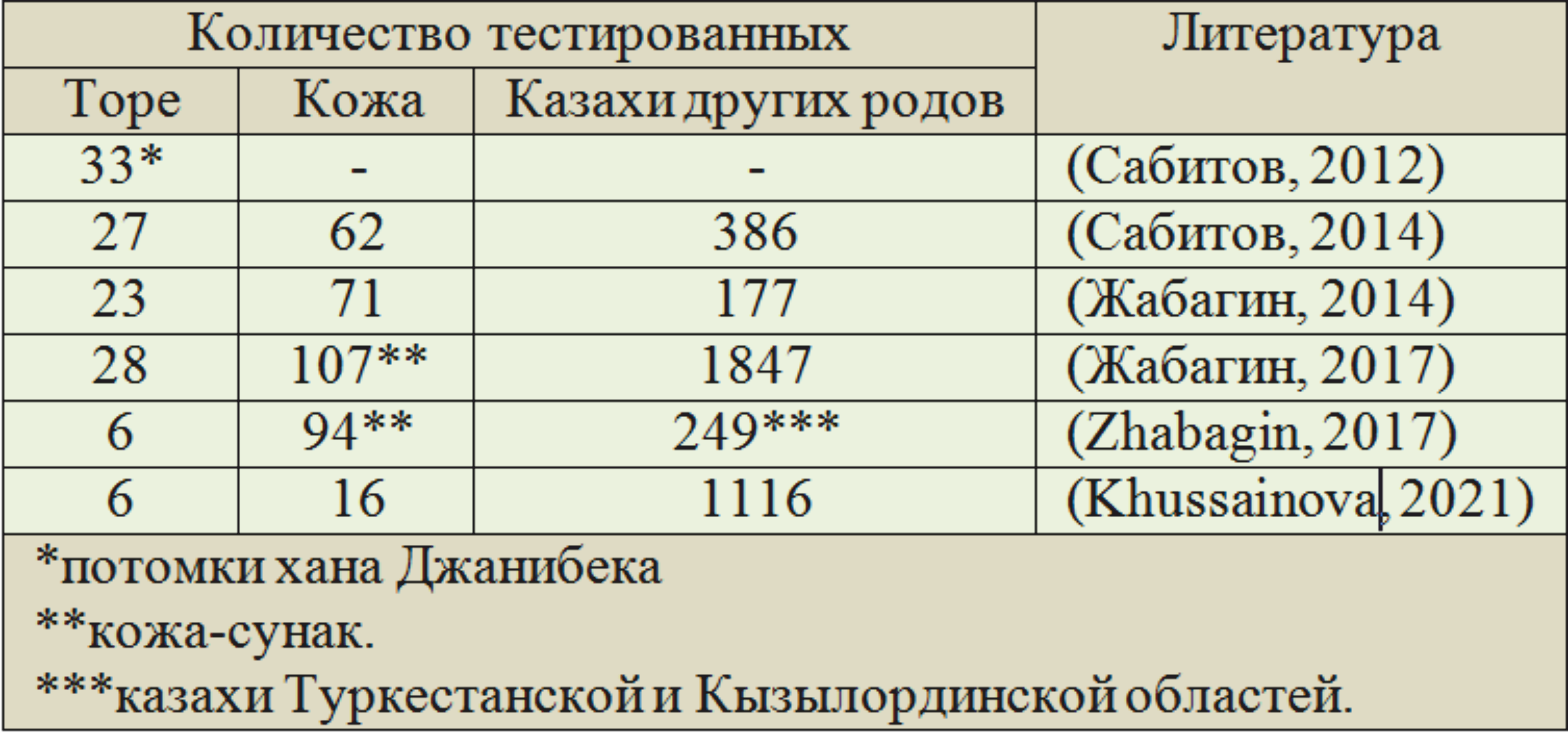
С данными по кожа имеется одна проблема. В публикации (Zhabagin, 2017) применяется два разных термина: «clans Kozha and Sunak» и «Kozha-Sunak tribe», в публикации (Khussainova, 2022) — «Kozha and Sunak clans» и «Kozha clan». Это при том, что в последней приведены результаты научной выборки, охватывающей всех казахов Казахстана.
Для анализа генетических портретов торе и кожа сформировано три массива гаплогрупп и их субкладов. Первый включает торе потомков Джанибека (Сабитов, 2012). Второй — данные по торе и кожа из публикаций (Жабагин, 2014, 2017; Сабитов, 2014; Khussainova, 2021), а также по казахским жузам (Khussainova, 2021). Данные по торе и кожа из публикации (Сабитов, 2014) рассмотрены отдельно. Приведённая в них номенклатура гаплогрупп и субкладов отличается от номенклатуры в других публикациях. Третий массив характеризует казахов Туркестанской и Кызылординской областей, а также узбеков Ташкента (Zhabagin, 2017). К нему добавлены данные по узбекам Туркестанской области (N=20) и таджикам Таджикистана (N=20) (Zhabagin, 2022). При учёте того, что в горной части Ташкентской области находится анклав таджиков, этот массив характеризует популяции Сырдарьинского региона: Ташкентской, Туркестанской и Кызылординской областей.
МЕТОДЫ
Методы анализа данных популяционной генетики основаны на этнической идентификации гаплогрупп Y-хромосомы и их субкладов. При этом она носит условный характер.
У монголов две субпопуляции — халха и ойраты (восточные и западные). Известны две крупные миграции монголов на запад за пределы территории их проживания в Центральной Азии. Первая — в процессе монгольских завоеваний в первой половине XIII в. В ней приняли участие халха и ойраты. Вторая — переход ойратов в Синьцзян, Семиречье и Северный Прикаспий в первой половине XVII в. По результатам первой миграции были сформированы улусы Джучи и Чигатая, второй — Калмыцкое ханство в Северном Прикаспии и Джунгарское в Семиречье и Синьцзяне. Монголы-ойраты, мигрировавшие в Северный Прикаспий, имеют этноним калмыки. Те, кто создал Джунгарское ханство (XVII в.–1759 г.), — калмаки. В русских письменных свидетельствах последних называют зюнгарами. В специальной литературе — джунгарами. Была и миграция калмаков (джунгар) с территории Джунгарского ханства в другие регионы после его разгрома войсками империи Цин в 1755–1759 гг. Одна их группа численностью 9 тыс. кибиток ушла в Ферганскую долину, другая (3 тыс. кибиток) — через Памир в Бадахшан (Афганистан). Ещё одна группа ( 2,5 тыс. Кибиток) мигрировала в пределы Калмыцкого ханства (Тюрин, 2024а).
Гаплогруппы Y-хромосомы C2, O и D называют монгольскими. По YTree гаплогрупп Y-хромосомы (YTree) нами выделено четыре субклада гаплогруппы C2, характерные для монголов: C2a1a2a-M48 «Калмыки» (по частотам доминируют у калмыков), C2a1a2b-M504 «Калмаки» (доминирует у калмаков), C2b-F1067 «Халха» (доминирует у монголов-халха) и C2-Y10418 «Авары» (этнически не идентифицирован). Такое распределение их частот позволяет выполнять этническую идентификацию носителей гаплогруппы C2. Работоспособность метода показана на популяциях Евразии.
1. Следы миграций ойратов в XVII и XVIII вв. выявлены в YTree (YTree) у татар Татарстана, Томской, Нижегородской и Рязанской областей (Тюрин, 2024б). Татары, носители гаплогруппы C2, являются их потомками. Потомков монголов-халха в Восточной Европе за исключением одного жителя Калмыкии и двух Чечни не имеется. Житель Калмыкии, скорее всего, калмык. У них по частотам доминирует субклад «Калмыки», но значимо представлен и субклад «Халха» (Тюрин, 2025а).
2. Выявлены следы миграции калмаков в XVIII в. Хазарейцы Афганистана и Пакистана являются их потомками (Тюрин, 2024а).
3. У родовых кланов тувинцев частоты гаплогруппы C2 составляют 4,4…43,5%. Доминируют её разные субклады — «Калмаки», «Калмыки» и «Халха». Для всех тувинцев структура гаплогруппы C2 по субкладам идентична её структуре у монголов (Тюрин, 2025б).
4. У казахов Казахстана 40–52 % носителей гаплогруппы C2. В этногенез казахов Старшего жуза определяющий вклад внесли калмаки, Младшего — калмыки. Их вклад в генетические портреты родов Среднего жуза дифференцированный. Монголов-халха на территории формирования этноса (восточная часть улуса Джучи и северо-восточная часть улуса Чигатая) не имелось. Частоты субклада «Халха» у них на уровне его частот у калмыков (Тюрин, 2025а). Исключение составляют
казахи рода канырат.
5. У сибирских татар имеются носители гаплогруппы C2. Резко доминирует субклад «Калмыки». Генетических следов экспансии халха в XIII в. у них не имеется (Тюрин, 2025в).
Ниже приведена привязка индексов субкладов гаплогруппы C2 к применяемой китайскими генетиками. «In Altaic-speaking populations, especially most Mongolicspeaking populations, there are four predominant paternal lineages of C2*-M217 clade, including C2b1a3a1-F3796 (previous C3*-Star Cluster), C2a1a1b-M48, C2b1a1a1a-M407 and C2a1a1a2a-F1756» (Wen, 2020). Субклад C2b1a3a1-F3796 непосредственно восходит к субкладу C2a1a2b-M504 «Калмаки» (Y-DNA), субклад C2a1a1b-M48 соответствует субкладу C2a1a2a-M48 «Калмыки», субклад C2b1a1a1a-M407 является дочерним субклада C2b-F1067 «Халха», субклад C2a1a1a2a-F1756 — субклада C2-Y10418 «Авары».
В приведенной цитате есть примечание: «C2b1a3a1-F3796 (previous C3*-Star Cluster)». При этом даётся ссылка на статью (Wei, 2018). Это давняя история. Авторы публикации (Zerjal, 2003) в гаплогрупе С*(×C3c) выделили кластер гаплотипов иназвали его C3*-Star Cluster. Оказалось, что его модальный гаплотип и гаплотипы, отстоящие от него на 1 и 2 мутации, имеют широкое распространение у носителей гаплогруппы C2. Предположили, что основателем кластера является Чингисхан. Тогда носители гаплотипов кластера — его прямые потомки. Их примерно 8 % азиатских мужчин (16 миллионов). Авторы публикации (Wei, 2018) эту гипотезу дезавуировали. C3*-Star Cluster является обычным для монголов. По нашей трактовке, относится к субкладу «Калмаки» (Тюрин, 2025а).
В соответствии с авторским вариантом филогении (Wei, 2018) C2*-Star Cluster гаплотипов относится к линии C2b1a3a1-F3796 субклада с маркером M504 (Рис. 1). Он является независимым от субклада с маркером M48. Также соотносятся эти субклады в проектах Y-DNA (Y-DNA) и YTree (YTree). В YTree в линии C2b1a3a1-F3796 доминируют жители Китая. Также она включает 7 хазарейцев из Афганистана и Пакистана, 1 калмыка и 3 татар из России, 7 жителей Казахстана (этнос указан для 4 — казахи), 4 монголов из Китая и 1 из России. На основе вышесказанного отредактировано обозначение субкладов гаплогруппы C2. Ниже они обозначаются C2-M48 «Калмыки» и C2-M504 «Калмаки».
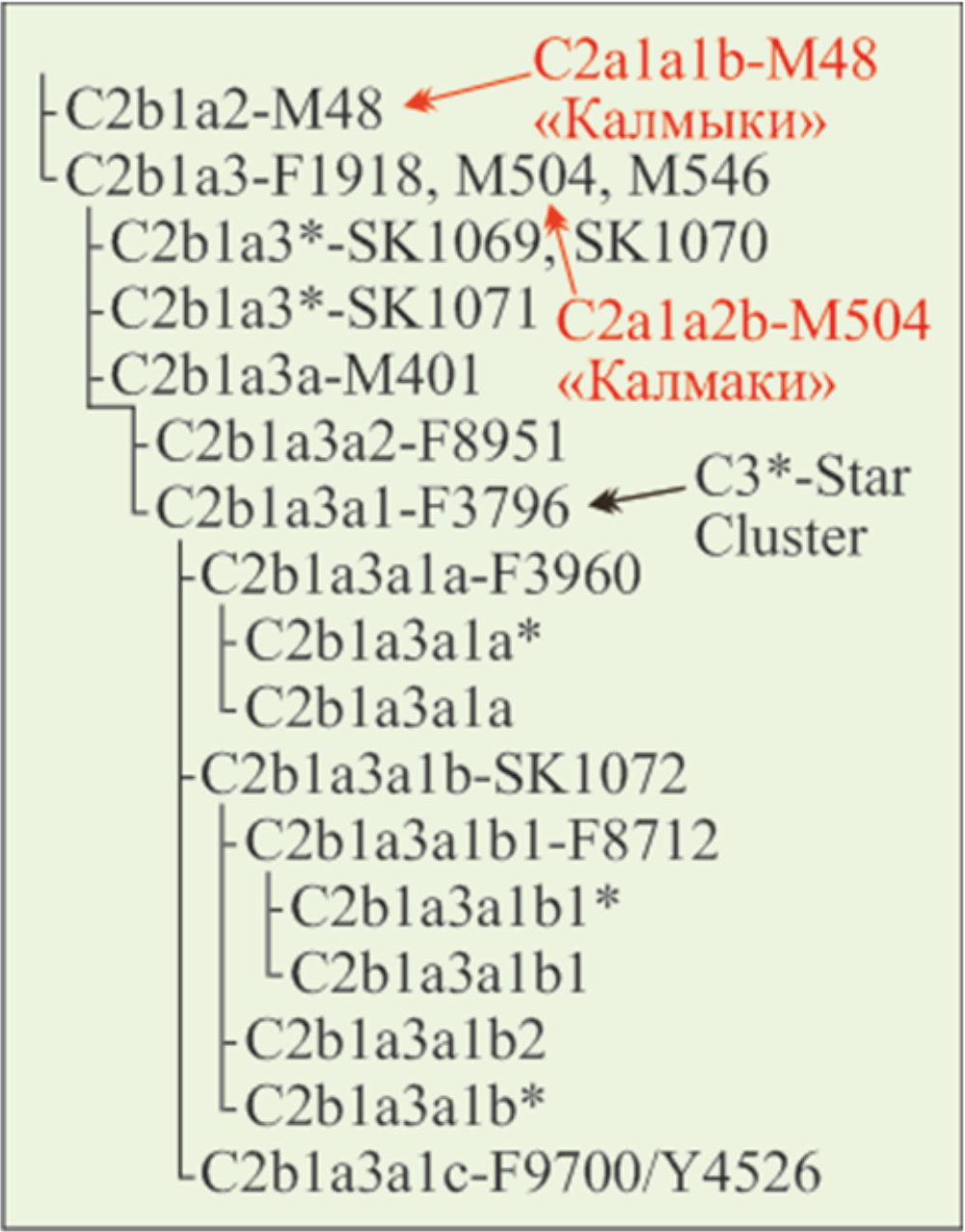
Рис. 1. Филогения Y-хромосомной линии C2*-Star Cluster (Wei, 2018). Информация красным цветом нанесена А.М. Тюриным
Анализ данных популяционной генетики, характеризующих торе и кожа, выполнен в два этапа. На первом рассмотрены только частоты в популяциях монгольских гаплогрупп Y-хромосомы и субклады гаплогруппы C2 — «монгольский» компонент. На втором — приведенные частоты других гаплогрупп и их субкладов (частоты за вычетом носителей монгольских гаплогрупп). В них выделены переднеазиатские, южные, степные и сибирские ассоциации гаплогрупп и их субкладов. В гаплогруппах R1a, R1b и N имеются европейские и азиатские субклады и линии.
В выборке по поволжским татарам они разделены (Акчурин, 2021). Но данные по казахам типизированы менее детально. Принято, что у них не имеется европейских линий гаплогрупп R1b и N. В гаплогруппе R1a выявлены только единичные носители славянской гаплогруппы R1a-M458. Скорее всего, в ней резко доминирует субклад R1a-Z93, называемый «степным».
В обосновании по фактическим данным нуждается только южная ассоциация гаплогрупп. Гаплогруппы J и G являются переднеазиатскими (Восточное Средиземноморье, Аравийский полуостров, Месопотамия и Кавказ). Но в регион проживания казахов они попали разными путями. Авторы публикации (Balanovsky, 2015) генотипировали гаплогруппу G1-M285 (N=5346, 27 евразийских популяций) и выполнили детальную филогенетическую реконструкцию. Идентифицировано пять ветвей гаплогруппы, по частотам доминирующих среди её носителей, у казахов, монголов, армян, башкир и жителей Индостана. Разнообразие гаплотипов гаплогруппы уменьшается от Западного Ирана до Центральной Азии. Сформулирована гипотеза: гаплогруппа G1-M285 перенесена в евразийские степи экспансией ираноговорящих народов. Авторы публикации (Zhabagin, 2017) отметили, что кожа-сунак и казахи рода аргын имеют родственные гаплотипы этой гаплогруппы. Её частоты у аргынов аномально высокие — 66,9 %. Гаплотипы гаплогруппы образуют кластер. Время жизни его основателя — 600±200 лет назад (Жабагин, 2016).
У уйгуров Хотана (N=478) 25,3 % носителей монгольских гаплогрупп и 15,7 % — гаплогруппы J. У уйгуров Турфана — 30,1 % и 12,6 % (Liu Shuhu, 2018). Приведённые частоты гаплогруппы J в двух регионах — 21,0 и 18,0 %. У двух других популяций Синьцзяна (кериян и лопур) её частоты — 28,2 % и 59,4 %. У них резко доминирует гаплогруппа J2 — 25,6 % и 57,8 %. У родов жалайыр (N=92) и дулат (N=92), в прошлом соседей уйгуров (населяли и район реки Или), значимо представлен субклад J2a-M401 — 14,1 % и 7,9 % соответственно (Аширбеков, 2018). Частоты гаплогруппы J у таджиков Таджикистана (N=40) — 15,0 %, персов восточного Ирана (N=62) — 44,2 % (Malyarchuk, 2012). У популяций Афганистана (N=212) — 13,2 % носителей гаплогруппы J2a (Haber, 2012). Носители гаплогруппы J1 у них не выявлены. Можно уверенно говорить, что предки торе и кожа, носители гаплогруппы J2, попали в регион проживания казахов с юга — Синьцзян, территории Таджикистана, Афганистана и восточного Ирана. Гаплогруппы R2, L и H1 являются специфичными для популяций Индии (Malyarchuk, 2012). Гаплогруппа T редкая. Её носители имеются и в Индии. У популяций Афганистана суммарная частота этих четырёх гаплогрупп составляет 18,9 % (Haber, 2012).
РЕЗУЛЬТАТЫ
У шести линий потомков хана Джанибека следующие частоты мажорных гаплогрупп: С3(×C3c) — 48,7 % (в основном линии Усека, Ондана и Адиля), R2a — 20,5 % (линия Ишима), R1a1a — 15,4 % (линия Самеке) (Сабитов, 2012). Непонятно, какие субклады включает гаплогруппа С3(×C3c). У почти половины потомков хана предками были ойраты или халха.
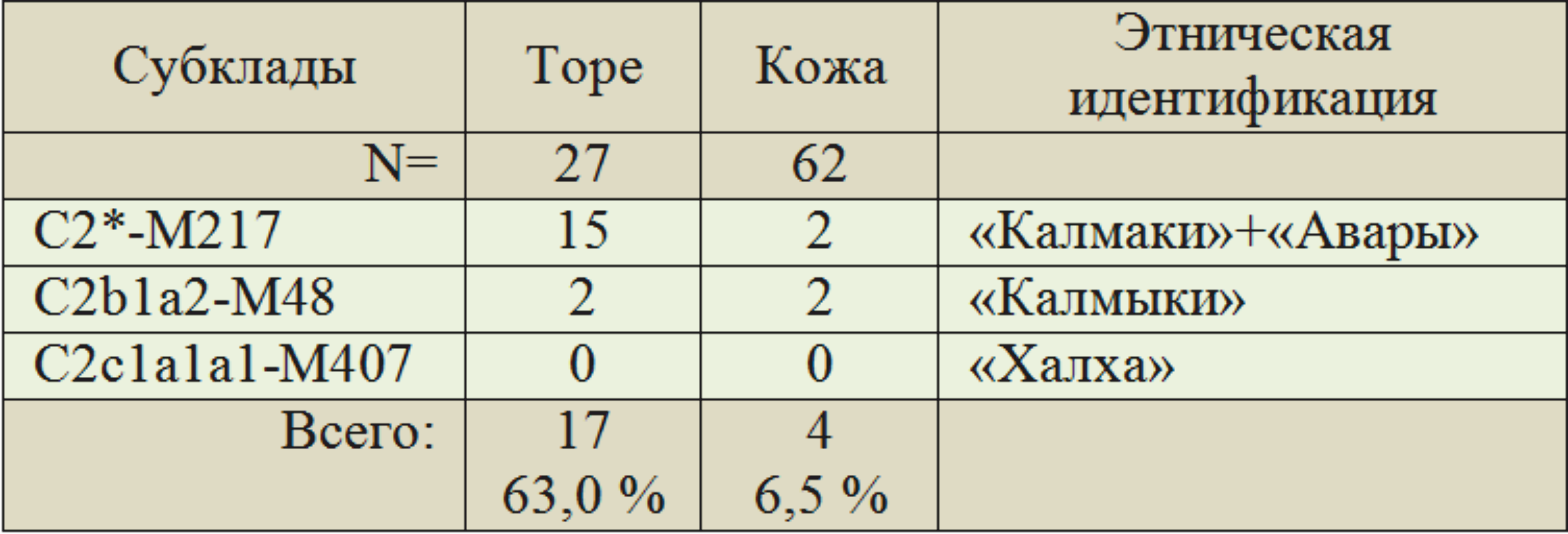
Частоты субкладов гаплогруппы C2 у торе и кожа из Казахского ДНК-проекта и наша их этническая идентификация приведены в таблице 2. У торе высокие частоты носителей гаплогруппы C2 — 63,0 %, у кожа их — всего 6,5 %. У казахов других 10 родов — 43,5 % (Сабитов, 2014). Среди торе и кожа нет потомков монголов-халха и минимальное число потомков калмыков. У коныратов (Средний жуз) (N=10) монголов-халха — 80,0 %. Это соответствует данным из публикации (Жабагин, 2017) (N=90) — 85,6 %. У других родов — 1,1 %.
Таблица 2. Структура субкладов гаплогруппы Y-хромосомы C2 (по количеству тестированных) у казахских родов торе и кожа в Казахском ДНК-проекте (Сабитов, 2014) и их этническая идентификация (А.М. Тюрин)
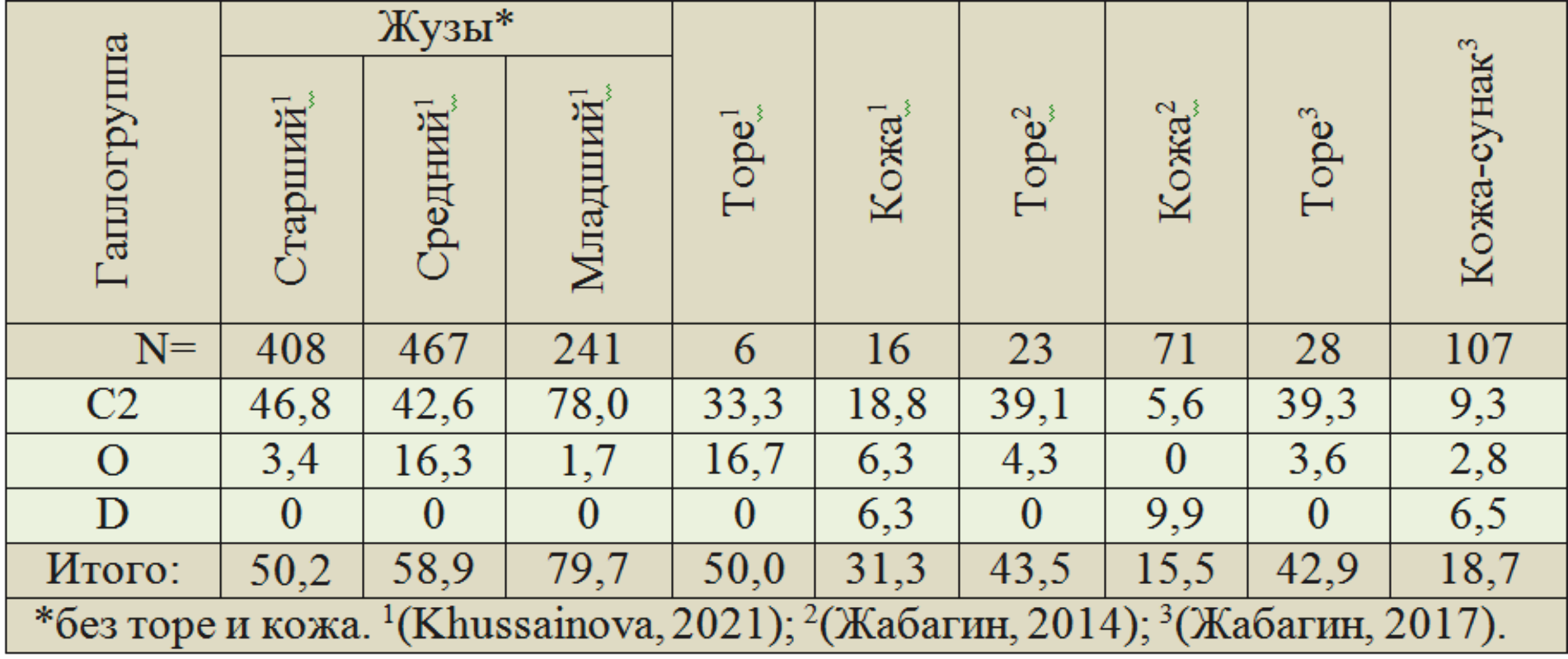
Частоты монгольских гаплогрупп выборок из публикаций (Жабагин, 2014, 2017; Khussainova, 2021) показаны в таблице 3. Для сопоставления приведены данные по трём жузам казахов (Khussainova, 2021). Частоты гаплогруппы C2 у казахов Старшего жуза — 46,8 %, Среднего — 42,6 %, Младшего — 78,0 %. У торе по трём выборкам 33,3…39,3 %, у кожа — 5,6…18,8 %. Частоты гаплогруппы O по жузам резко дифференцированы. Носители гаплогруппы D у казахов жузов и торе не выявлены. У кожа их — 6,3…9,9 %.
Таблица 3. Структура монгольских гаплогрупп Y-хромосомы (%) у казахов
В выборке из публикации (Жабагин, 2014) в гаплогруппе C2 выделено два субклада — C3c-M48 и C3(×C3c)-M217(×M48). У торе — 4,3 % и 34,8 % их носителей. Гаплогруппа C3c-M48 идентифицируется однозначно — «Калмыки». Гаплогруппа C3(×C3c)-M217(×M48) по частотам доминирует у казахов родов жалайыр (38,0 %) и керей (64,9 %), а также у монголов рода борджигин (34,3 %). Частоты гаплогруппы C3c-M48 у трёх родов — 2,0 %, 10,4 % и 18,2 % соответственно. В другой выборке у кереев (N=207) по частотам доминируют носители гаплогруппы C2 — 85,8 % (Zhabagin, 2024). Структура её субкладов: «Авары» —2,9 %, «Калмаки» — 79,6 %, «Калмыки» — 1,9 %, «Халха» — 1,4 % (Тюрин, 2025а). У жалайыров (N=103) всего один носитель гаплогруппы C2-M407 (Жабагин, 2014). Таким образом, казахские кереи и жалайыры не восходят к древнемонгольским родам. Их предки были ойратами (калмаками).
В филогенетической сети гаплотипов (по 17 STR-маркерам) гаплогруппы C3(×C3c)-M217(×M48) в популяциях Евразии (N=783) обособилось три кластера (Жабагин, 2014). Кластер α (дата общего предка 1000±300 лет назад) включает предполагаемый гаплотип Чингисхана, то есть является C3*-Star Cluster. В него попали и гаплотипы торе. Их предками были калмаки. Кластер γ (1100±400 лет назад) сформирован гаплотипами рода конырат. Это единственный род казахов, предками которых были халха. Кластер β (600±300 лет назад) включает только монголов. Данных для его этнической идентификации недостаточно. В выборке из публикации (Жабагин, 2017) у торе и кожа носителей субклада C2-M407 не имеется. Носителей субклада C2-M48 — 3,6 % и 2,8 % соответственно. В выборке из публикации (Khussainova, 2021) субклады в гаплогруппе C2 не выделены.
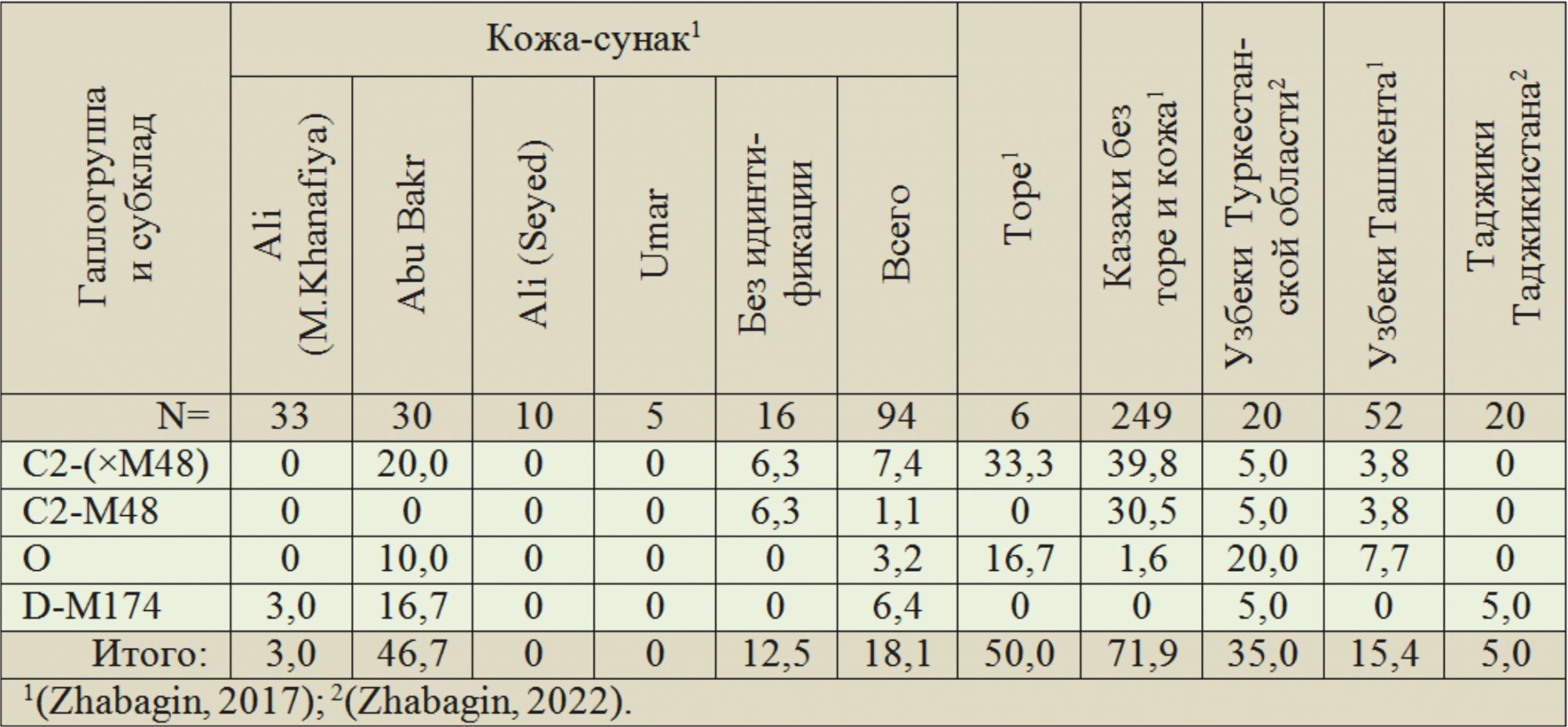
Данные по популяциям Сырдарьинского региона приведены в таблице 4. В клане кожа-сунак высокий процент носителей монгольских гаплогрупп у Abu Bakr — 46,7 %. У остальных суммарно — 4,7 %. У казахов региона — 71,9 %. Всего у кожа-сунак их частоты — 18,1 %, примерно находятся на уровне этого параметра у узбеков Ташкента — 15,4 %, но в два раза ниже, чем у узбеков Туркестанской области, — 35,0 %. У узбеков обоих регионов соотношение частот гаплогрупп O и C2 — 2,0 и 1,0. У казахов — 0,02.
Таблица 4. Структура монгольских гаплогрупп Y-хромосомы (%) у казахов Туркестанской и Кызылординской областей в сопоставлении с узбеками и таджиками
В выборке из публикации (Сабитов, 2014) у торе высокие частоты гаплогруппы R2 — 22,2 %. Частоты гаплогрупп G1, J и Ra1 — 3,7 %. У кожа — R1a — 27,4 %, J — 12,9 % и R1b — 11,3 % (в том числе R1b1a2-M269 — 9,7 %). У этих двух родов носители гаплогруппы N не выявлены. У других казахских родов 3,6 % их носителей. Носители 20 гаплогрупп кожа в статье не приведены. Субклад R1b1a2-M269 в YTree (YTree) включает представителей Западной Европы, Северного Кавказа и арабов. По частоте резко доминирует у башкир рода бурзян (Юсупов, 2023). Скорее всего, гаплогруппа R1a представлена в основном субкладом R1a1-Z93. С высокими частотами он распространён у популяций Средней Азии, Алтая, Афганистана, Индии и других регионов Евразии.
Приведённые частоты не монгольских гаплогрупп по всем казахам показаны в таблице 5. Выборка, включающая 3 торе, в рассмотрение не включена.
Таблица 5. Структура не монгольских гаплогрупп Y-хромосомы (%*) у казахов
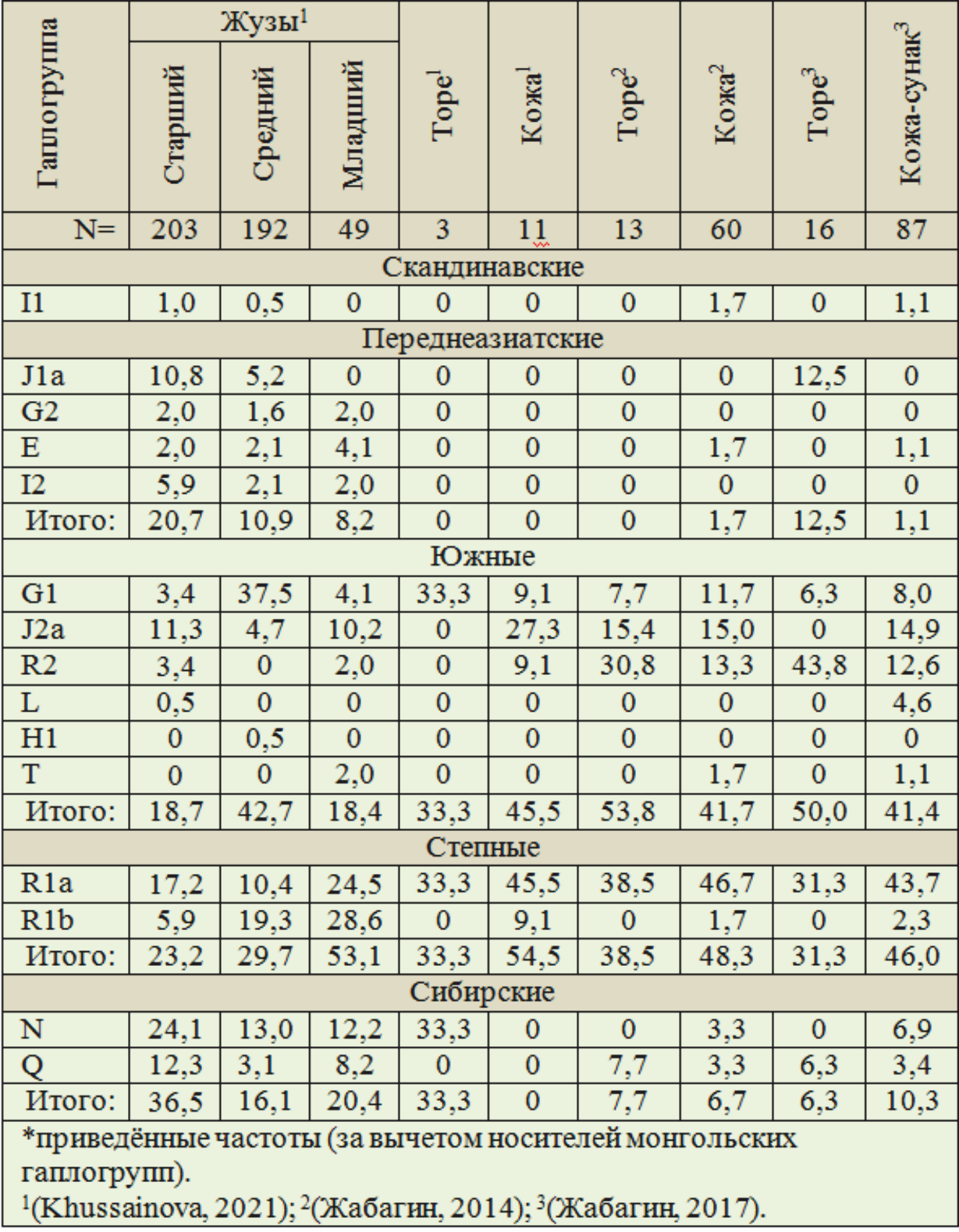
1. У казахов по жузам 8,2…20,7 % носителей переднеазиатских гаплогрупп. У торе и кожа их практически не имеется. Только в одной из двух рассматриваемых выборок торе есть носители переднеазиатской гаплогруппы J1a — 12,5 %.
2. Частоты сибирских гаплогрупп у торе и кожа — 6,3…10,3 %. У казахов их в разы больше — 16,1…39,5 %.
3. По степным гаплогруппам соотношение казахов и аристократических родов дифференцированно. У первых значимо представлена гаплогруппа R1b — 5,9…28,6 %., у торе её носителей не имеется, у кожа — 1,7…9,1 %. Гаплогруппа R1a по частотам доминирует у аристократических родов — 31,3…46,7 %, у казахов — 10,4…24,5 %.
4. Суммарные частоты южных гаплогрупп: у торе — 50,0 и 53,8 %, у кожа — 41,4…45,5 %, степной: у торе — 31,3 и 38,5 %, у кожа — 46,0…54,5 %. У торе несколько выше, чем у кожа частоты южных гаплогрупп, но ниже степных. Суммарно частоты южных и степных гаплогрупп у торе — 81,3 и 92,5 %, у кожа — 87,4…100 %. Кожа из научной выборки (Khussainova, 2021) являются носителями исключительно
гаплогрупп этих ассоциаций.
5. Близкие частоты у торе и кожа отмечаются по гаплогруппам G1 — 6,3…11,7 %, R1a — 31,3…46,7 % (у торе несколько ниже, чем у кожа), R1b — 0…9,1 % и суммарно по L, H1 и T (у торе их носители не имеются, у кожа они единичные). У торе частоты гаплогруппы R2 — 30,8 и 43,8 %, в разы выше, чем у кожа — 9,1…13,3 %. 6. В одной из двух выборок торе 15,4 % носителей гаплогруппы J2a, в другой
они не выявлены. У кожа — 14,9…27,3 %.
Распределение приведённых частот немонгольских гаплогрупп у популяций Сырдарьинского региона имеет яркие особенности (Табл. 6). Выборка, включающая 3 торе, в рассмотрение тоже не включена.
Таблица 6. Структура не монгольских гаплогрупп Y-хромосомы (%*) у казахов Туркестанской и Кызылординской областей в сопоставлении с узбеками и таджиками
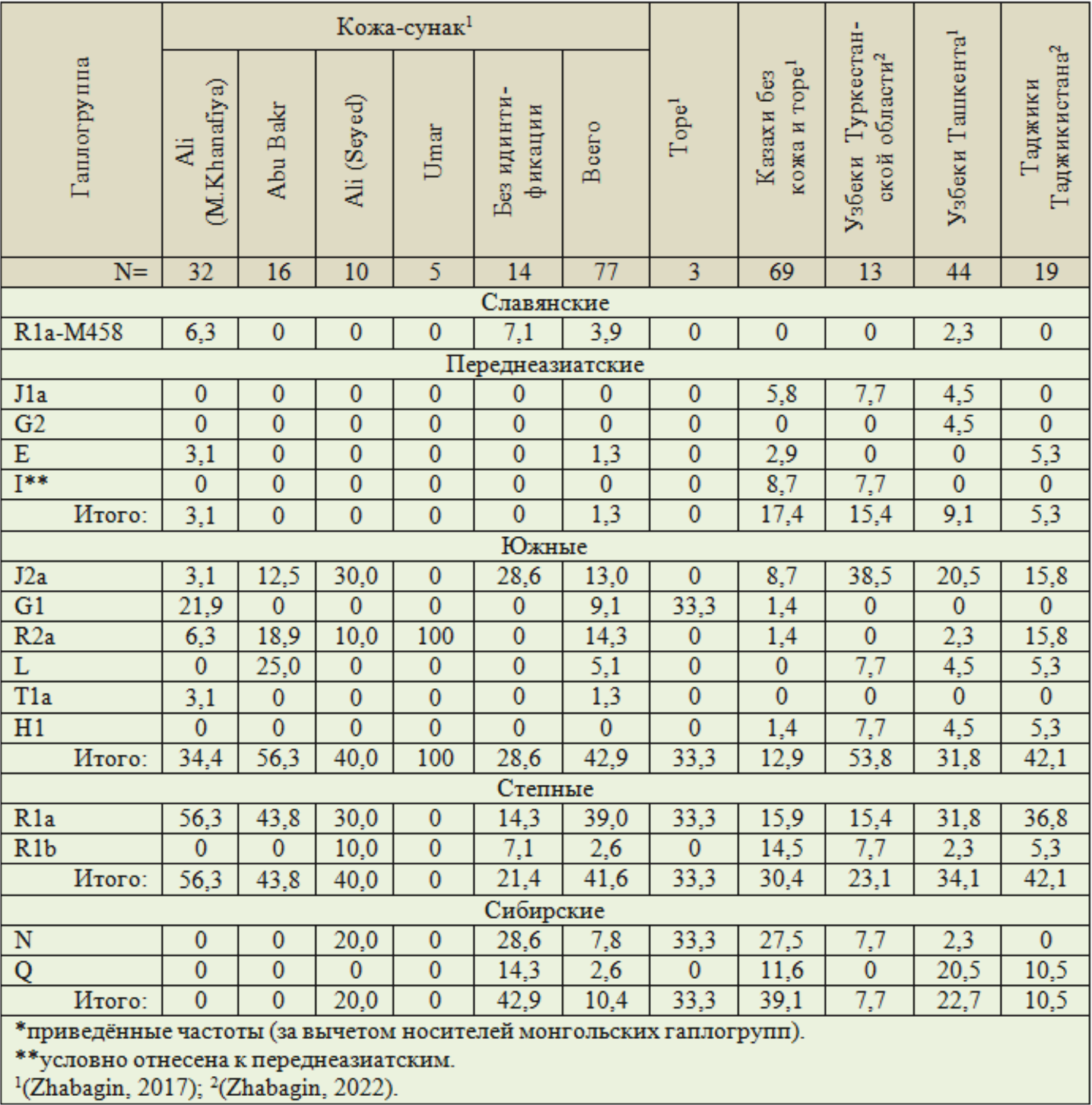
1. У кожа-сунак имеются три носителя славянского субклада R1a-M458, один его носитель у узбеков Ташкента. Субклад имеет две ветви — центрально-европейскую и западнославянскую (Рожанский, 2018). Скорее всего, этот субклад попал к отмеченным популяциям от славян переселенцев или «русского полона». Влияние последнего на этногенез поволжских татар рассмотрено в публикации (Тюрин, 2025б).
2. Суммарные частоты переднеазиатских гаплогрупп формируют «клин»: казахи — 17,4 %, узбеки Туркестанской области — 15,4 %, узбеки Ташкента — 9,1 %, таджики — 5,3 %, кожа-сунак — 1,3 %.
3. У казахов и узбеков имеются носители гаплогруппы J1a — 4,5…7,7 %, у кожа-сунак они не выявлены.
4. У кожа-сунак и таджиков Таджикистана совпадают суммарные частоты южных гаплогрупп — 42,9 и 42,1 %. Практически совпадают частоты четырёх гаплогрупп: J2a — 13,0 и 15,8 %, R2a — 14,3 и 15,8 %, L — 5,1 и 5,3 % и T — 1,3 и 0 %. Всего —33,7 и 36,9 %. Наличие у узбеков и таджиков носителей гаплогруппы H1 скорее всего связано с проживанием среди них цыган люли, выходцев из Индии. У кожа-сунак 9,1 % носителей гапогруппы G1. У узбеков и таджиков они не выявлены. У казахов представлены минимально — 1,4 %. Ситуация по этой гаплогруппе рассмотрена выше. Она могла попасть к кожа-сунак с территории Ирана. Могла попасть и от аргынов.
5. Отмеченная закономерность в пункте 4 проявилась и в двух степных гаплогруппах. Их частоты у кожа-сунак и таджиков — 41,6 и 42,1 %. Частоты гаплогруппы R1a — 39,0 и 36,8 %, R1b — 2,6 и 5,3 %. Немного ниже частоты этих гаплогрупп у узбеков Ташкента — 31,8 и 2,3 %.
6. Частоты сибирской гаплогруппы N тоже формируют «клин»: казахи — 27,5 %, узбеки Туркестанской области — 7,7 %, узбеки Ташкента — 2,3 %, таджики — 0 %. У кожа-сунак частоты этой гаплогруппы — 7,8 %, на уровне узбеков Туркестанской области.
7. Гаплогруппа Q значимо представлена у казахов — 11,6 %, узбеков Ташкента — 20,5 % и таджиков — 10,5. У узбеков Туркестанской области не выявлена. У кожа-сунак представлена минимально — 2,6 %. Гаплогруппа Q считается сибирской. Её частоты у кетов и селькупов 94 % и 66 %. Высокие частоты гаплогруппы у северных алтайцев. Есть ещё одна популяция с высокой долей носителей гаплогруппы Q. Это туркмены, проживающие на территории исторического региона Трансоксиана (Каракалпакстан). У них (N=83) — 73,5 % носителей этой гаплогруппы (Схаляхо, 2016; Zhabagin, 2017).
8. В южной ассоциации гапогрупп имеется ещё один «клин». Частоты гаплогруппы R2a у таджиков — 15,8 %, узбеков Ташкента — 2,3, узбеков Туркестанской области — 0 %. Если эту гаплогруппу исключить, то суммарные частоты двух ассоциаций — южной и степной, будут примерно одинаковыми: узбеки Туркестанской области — 76,9 %, узбеки Ташкента — 63,6 %, таджики — 68,4 %. У узбеков двух объединённых выборок — 66,7 %.
9. У узбеков Ташкента и Туркестанской области соотношение частот гаплогрупп O и C2 не характерно для казахов Сырдарьинского региона и монголов (Дамба, 2018). Гаплогруппа O по частотам доминирует у китайцев. Можно уверенно предполагать, что джунгарский ясырь включал и китайских полоняников. Часть узбеков, носителей гаплогруппы O, является их потомками.
Формальные выводы.
1. Число носителей монгольских гаплогрупп у торе — 42,9–50,0 %, находится на уровне их носителей у казахов Старшего жуза — 50,2 %, но несколько ниже, чем у них. У кожа в двух больших выборках число носителей этих гаплогрупп — 15,5 и 18,7 %, резко занижено относительно казахов. Среди торе, носителей монгольской гаплогруппы C2, не имеется потомков халха, минимально представлены потомки калмыков. Основная масса их предков была калмаками и носителями не идентифицированного субклада «Авары».
2. Генетические портреты торе и кожа по немонгольским гаплогруппам принципиально идентичны. У них резко доминируют носители южной и степной ассоциаций гаплогрупп. Отмечаются расхождения в частотах гаплогрупп R2 и J2a. Частоты первой выше у торе, а второй — у кожа. Носители переднеазиатской и сибирской ассоциации представлены минимально. Этим они отличаются от казахов (в среднем) других родов.
3. Генетические портреты кожа-сунак, таджиков Таджикистана и узбеков двух выборок принципиально идентичны. Ассоциация их гаплогрупп названа «таджики».
ОБСУЖДЕНИЕ
В восточной горной части Ташкентской области находится анклав таджиков (посёлки Бурчмулла, Сиджак, Богустан, Нанай и др.). В прошлом они проживали и в долине реки Ангрен. Важная информация об их предках по состоянию на 1899 г. приведена в публикации (Кузнецов, 1900). К ней даны наши комментарии.
1. До прихода русских официальным языком в регионе являлся персидский, на котором пишут таджики. Таджики говорят на таджикском и узбекском языках. Автор описал процесс их перехода в сарты. Представляется, что он застал завершение этого процесса на равнине и в предгорьях. При учёте принципиальной идентичности генетических портретов узбеков Сырдарьинского региона и таджиков Таджикистана можно сделать вывод: в прошлом здесь проживали представители одной популяции. Скорее всего, они же проживали в городах Ферганской долины, а также междуречья Сырдарьи и Амударьи. Из этой же популяции сформировались рода торе и кожа.
2. У таджиков было большое количество святых, могилы которых имеются на мазарах. Таджики считали их арабами выходцами из Мекки или Медины. Причём все святые современники друг другу. Это и есть первая генерация проповедников ислама. Потомки большинства из них стали ходжами.
3. Во время уборки урожая в регион на подработку прибывают казахи с Сырдарьи и Келеса. Автор привёл названия их родов. В том числе и один род ходжа, считающий себя потомками первых халифов.
4. Женщины у таджиков пользуются большой свободой, вплоть до того, что «пошаливают». Адюльтер не редкость.
Всего выделяется 17 кланов кожа, включая сунак (Маликов, 2014). Автор публикации привёл этнографические данные по трём кланам ходжей Южного Казахстана: хорасан-ходжей, кылышты-ходжей и узбекоязычных ходжей. До национального размежевания в регионе (с 1924 г.) ходжи различали друг друга только по генеалогическому принципу, после стал доминировать этнический критерий, основанный на языковой
аффилиации. То есть, в Сырдарьинском регионе первоначально единое сообщество хаджей в советский период разделилось на казахских кожа и узбекских ходжей.
Хорасан-ходжи в досоветский период кочевали. Считают себя потомками халифа Али. Вероятно, получили свое название от исторической провинции Хорасан. Проживали в Узгенте, Тугискене, Джанакургане, Кызыл-Орде. Кылышты-ходжи относяся к сейид-ходжам через двух святых. Информаторы утверждают, что родина кылышты-ходжей в Узгенте (Маликов, 2014). Этот город был расположен на территории Жанакурганского района западнее Сырдарьи. От него сохранилось городище (Городище). Тугискент — село и одноимённый могильник в Жанакурганском районе.
Узбекоязычные ходжи идентифицируют себя как узбеки (Маликов, 2014). Проживают в Туркестане и других городах Южного Казахстана. В Туркестане представлены потомками суфийских святых Гаухар-она, Али-ходжи, Садр-ота и др. По преданию, некоторые из них являются потомками ходжей из Бухары. По другому преданию, они потомки людей, служивших у суфийского лидера Ходжи Ахмеда Яссеви.
Этнографические сведения по рассматриваемому вопросу приведены в публикации (Ахметова, 2013). В XVIII в. кожа, потомки миссионеров шиитов, прибыли из Туркестана вслед за Аблаем на территорию бывшей Кокчетавской области. «Здесь они открыли мечети и мектебы, обучая местных казахов арабскому языку и Корану. Затем перевезли из Туркестана своих родственников. Кочуя с казахскими родами,
они дошли до своего нынешнего места проживания в Омской области» (Ахметова, 2013: 13). Сегодня «на кладбище с одной стороны покоятся останки потомков шиитов, с другой стороны потомков суннитов» (Ахметова, 2013: 14). Эта информация подтверждает предположение автора публикации (Маликов, 2014) о том, что предки хоросан-ходжей были выходцами из Хоросана. На то, что они были шиитами, указывает их генеалогическая легенда. Халиф Али почитается шиитами. Имеются сведения и о наличии на территории Среднего жуза кожа, считающих себя потомками кашгарского святого Аппакходжи (Бисултанова, 2017).
Перовский и Казалинский уезды Сырдарьинской области занимали нижнее течение реки Сырдарьи. По состоянию на 1863 г. на территории Казалинского уезда было 525 к. (к. — кибитка) (3,0 %) ходжей, в округе Перовска — 700 к. (3,0 %) (Радлов, 1989). В данных Туркестанского Статистического комитета за 1870 г. различаются ходжи и сунак (Картаева, 2010). В Яныкурганской волости Перовского уезда среди других родов казахов отмечены сунаки (149 к.) и ходжи (92 к.), что в сумме составляет 7,8 % от общего числа кибиток. В Кентубской и Кармакчинской волостях отмечены только ходжи — 432 и 10 к., 20,8 и 0,6 %. В данных по другим семи волостям эти рода не упоминаются. Автор публикации (Алпысбес, 2006) привёл список кланов кожей казахов. Всего 17. Сунак среди них не значится.
Всего ходжей в Перовском уезде (по числу кибиток) — 2,7 %, сунак — 0,7 %. В Казалинском уезде выделен особый род, состоящий из торе, толенгутов и ходжей. Их 4,0 %. Теленгуты — это казахский род, не входящий в систему жузов (как торе и кожа). По записям офицеров генерального штаба автор публикации (Картаева, 2010) локализовала регион проживания ходжей — по Сырдарье и Куандарье, около крепости Камыскурган. Здесь, скорее всего, имеется ввиду укрепление Кокандского ханства Камыш-Курган, которое находилось севернее Ак-Мечети (Кызылорды) (Турсунов, 2019). Для сравнения — по переписи 1861 г. в Каркалинском округе Западно-Сибирского генерал-губернаторства (территория Среднего жуза), ходжей насчитывалось 206 человек, 0,3% от всего населения (Бисултанова, 2017).
Сунаки в 1870 г. были локализованы в Яныкурганской волости. Из 77 тестированных кожа-сунак 65 — из Жаныкурганского района (Zhabagin, 2017). По другой информации, формирование субэтнической группы сунак связано с жителями Сыгнака (Сандыбаева, 2025). Сунак и Сыгнак — это два названия древнего города, развалины которого — Сунак-Курган — находятся в Жаныкурганском районе.
Сыгнак находился на правом берегу Сырдарьи. С XIV в. был политическим, военным и торгово-экономическим центром на границе Дешт-и Кипчак и земледельческого региона. Им владели правители левого крыла улуса Джучи (Акимбеков, 2021) Со второй половины XV до конца XVI вв. Сыгнак был столицей Казахского ханства. Это превратило его в город-мавзолей (Нагаминэ, 2020). В последней публикации приведено свидетельство о начале XVI в.: «Могилы и гробницы всех ханов узбеков из шибановцев, юрты которых в старину были близки к Туркестану [Сырдарьинский регион], находятся в самом Сыгнаке и в его округе, потому что он является окраиной благоустроенной области […] Поневоле, из именитых ханов Дашта гроб каждого, кому наступал час предопределенной смерти, обязательно доставляли в Сыгнак, над могилой его воздвигали здание, похожее на купол» (Нагаминэ, 2020: 543). Сыгнак являлся и центром земледельческого района. Сельхозугодья вокруг него орошались двумя каналами (Мургабаев, 2022). Один был выведен из Сырдарьи, другой — из озера, заполняемого во время её весенних разливов. Использовалась и вода небольших речек, стекающих с гор Каратау. Имеются письменные свидетельства, выданные религиозным деятелям города, о том, что они имеют право возделывать землю, используя воды конкретных арыков и речек.
Последние упоминания о Сыгнаке в документах первой половины XVII в.: рядом с Сыгнаком находился город Узгент, родина кылышты-ходжей. Это один из путей формирования кланов кожа. В данном случае два клана сформировались внутри городов. С прекращением их существования стали кочевниками. Но возможно и то, что первоначально кочевниками стала часть горожан, включая религиозных деятелей и небольшое количество кожа. Позднее бывшие жители Сыгнака стали сунаками, ещё позднее — кожа-сунаками. Главное здесь то, что эти кланы сформировались из жителей городов Сырдарьинского региона, а они являлись частью популяции «таджики».
Казахами торе-чингизидами признавались потомки ханов Жанибека (династия тука-тимуридов 13-го сына Джучи) и Шибана (5-й сын Джучи) (Кият, 2024). Только они могли быть казахскими ханами (Акимбеков, 2021). Но в регионе были и торе, которые не являлись чингизидами. Среди ходжей был клан, члены которого возводили свою родословную к Пророку Мухаммеду. Себя они называли «тура». Титул «тура» или «тора» носили и ханские сыновья. В Бухарском эмирате вплоть до XX в. наследника престола называли тура. Этим словом называли и знатного человека, предводителя военного отряда. Мусульманские историки в XVIII в. называли «тура» правителей Джунгарского ханства. В данном случае «тура» — титул, синоним титула «хан» (Гафуров, 1987).
А. Гафуров считает слово «торе» тюркским (1987: 94). Но основания не приведены. В древнетюркском словаре этого слова нет (Древнетюркский…,1969). Ближайшее по значению TÖRÄ — «закон, обычай». Перевод имени Дара(Därä) с древнеиранского — «обладатель доброй, благой власти» (Гафуров, 1987).В Wikipedia приведено 12 статей о правителях с именем Дарий. Оно образовано от древнеперсидского dar «владеть». Скорее всего, в большинстве письменных свидетельств указан титул правителя. Другая форма DÄRÄ — ТОРЕ (Д>Т). Общий перевод — «властелин». Торе является частью слова тархан (торе+хан). Это титул, т.е. титул торе, применяемый у казахов к чингизидам, не является уникальным.
По данным популяционной генетики по казахским торе не представляется возможным сформулировать обоснованную гипотезу о гаплогруппе Чингисхана. Две гипотезы приведены ниже.
Пока не найдено ни одного захоронения членов семьи Чингисхана, включая его самого (Lkhagvasuren, 2016). Археологами предполагается, что таковыми являются пять погребений могильника Таван Толгой (Восточная Монголия). У останков трёх мужчин определена гаплогруппа R1b-M343. Сегодня она с высокими частотами распространена в Западной Европе. Заметно представлена на её других территориях и в сопредельных регионах Азии. Авторы отмеченной публикации предположили, что Чингисхан был её носителем. Но у казахских торе носители этой гаплогруппы не выявлены.
Представители клана Лу из Северо-Западного Китая утверждают, что являются потомками Тогана, шестого сына Чингисхана. Выполнены генетические исследования (N=30) (Wen, 2019). Все тестированные являются носителями монгольских гаплогрупп. По частоте доминирует субклад C2b1a1b1-F1756 — 53,3 %. По мнению авторов публикации, она может быть ещё одним кандидатом на гаплогруппу Чингисхана. По нашей трактовке, это субклад «Авары».
В публикации (Neparaczki, 2019) приведены результаты тестирования останков из погребений на территории Венгрии гуннов (V в.), аваров (конец VI–VII вв.) и мадьяр (конец IX–X вв.). По двум образцам из погребений аварского периода определена гаплогруппа С2. В публикации (Maroti, 2022) даны результаты тестирования останков из погребений этих же периодов. Гаплогруппа С2 определена в одном образце погребения аварского и в одном — мадьярского периодов. Результаты тестирования приведены в YTree (YTree). ПалеоДНК попали в субклад C-Y10418. На этом основании он назван «Авары» (Тюрин, 2024б). Недавно в этот субклад добавлены 43 палеДНК с территории Австрии, объединенные в 5 линий. Представляется целесообразным выполнение более глубокого генотипирования гаплогруппы C2 казахских торе. В отмеченных публикациях субклады C2-M504 «Калмаки» и C2-Y10418 «Авары» не разделены. У монголов (N=852) 58,1 % носителей гаплогруппы C2 (Дамба, 2018). Носителей субклада «Авары» среди них всего 6,9 % (Тюрин, 2025а).
Авторы публикации (Сабитов, 2012) считают, что адюльтер является единственным механизмом инкорпорации в генетический портрет чингизидов чуждых им гаплогрупп. Это не исключается. Но был и другой механизм — шежире. «Если опираться на все вышеописанные шежире султанов, в Казахстане оказались разными судьбами чингизиды из домов Сибирского, Казанского, Хивинского ханств, Аральского и Каракалпакского владений, Ногайского юрта. Потомки всех этих султанов на данный момент проживают в Казахстане» (Кият, 2024: 1519). Иными словами, авторы статьи считают шежире вполне достоверными документами.
Другое мнение по шежире у автора публикации (Алпысбес, 2006): Ходжи «создавая, свою генеалогическую историю, не ограничивались этим и участвовали в создании, закреплении генеалогических историй чингизовых династий и, как показывает шежире казахов, внесли свою определённую лепту в формирование структуры генеалогических систем казахских родов и родовых объединений» (2006: 306). По-другому и быть не могло. Молодому Казахскому государству нужен был этнический миф. И он был создан. Шежире потомков Чингисхана обзавелись те, кому это было нужно и кто мог их заказать специалистам. Родовая знать была на виду, и сделать это для неё было проблематичным. А кожа и жители городов Сырдарьинского региона смогли. Поэтому торе и кожа практически не отличимы по ассоциациям немонгольских гаплогрупп. Соответствующими шежире смогли обзавестись и калмаки, потерявшие связь со своими родами. Их интересовал титул торе. Поэтому у торе носителей монгольских гаплогрупп в разы больше, чем у кожа.
ЗАКЛЮЧЕНИЕ
1. В генетических портретах кожа и торе выделено два этнических компонента — «таджики» и «монголы». Популяция «таджики» в прошлом занимала территории Таджикистана, Северного Афганистана, Восточного Ирана. Проживала в городах Сырдарьинского региона, возможно, Ферганской долины, а также междуречья Сырдарьи и Амударьи. «Монголы» маркируются гаплогруппами Y-хромосомы C2, O и D.
2. У торе частоты монгольских гаплогрупп в разы выше, чем у кожа. Среди носителей субкладов гаплогруппы C2 не имеется потомков монголов халха. Минимально представлены потомки калмыков. Большинство являются потомками калмаков и носителей этнически не идентифицированного субклада «Авары».
3. Предки-арабы с Аравийского полуострова у кожа не выявлены.
4. Данные популяционной генетики по торе и кожа не соответствуют их генеалогическим легендам.
5. Предполагается, что большая часть шежире кожа и торе составлена в рамках формирования этнического мифа Казахского государства. В большинстве из них указана нереальная родословная семей.
ЛИТЕРАТУРА
Акимбеков, С.М. (2021). История степей. Феномен государства Чингисхана в истории Евразии. 4-е издание. Алматы: ТОО «Институт Азиатских исследований».
Акчурин, М.М., Владимиров, О.О., Салихов, Р.Р., Хакимов, Р.С. (2021). Генофонд татар: историко-генетическое исследование. Гаплогруппы Y-хромосомы. Казань. Институт истории им. Ш. Марджани АН РТ.
Алпысбес, М. (2006). Ходжи в генеалогической системе казахского народа. Парасат парызы. Қарағанды: ҚарМУ баспасы. 303–307.
Ахметова, Ш.К. (2013). Қожа в этнической истории казахов Западной Сибири. Казанская наука. 4. 12–14.
Аширбеков, Е.Е., Хрунин, А.В., Ботбаев, Д.М. и др. (2018). Молекулярно-генетический анализ популяционной структуры казахского племенного объединения Старший жуз на основе полиморфизма Y-хромосомы. Молекулярная генетика, микробиология и вирусология. 36. 2. 72–75.
Бисултанова, З.С.М., Горбунова, С.В. (2017). Ходжи в традиционном казахском обществе. Актуальные проблемы истории, документоведения и педагогики. 89–91.
Гафуров, А. (1987). Имя и история. Об именах арабов, персов, таджиков, тюрков. Словарь. М.: «Наука».
Городище Озгент. Древние городища Кызылординской области. https://silkadv.com/en/content/gorodishche-ozgent
Дамба, Л.Д., Балановская, Е.В., Жабагин, М.К. и др. (2018). Оценка вклада монгольской экспансии в генофонд тувинцев. Вавиловский журнал генетики и селекции. 22. 5. 611–619.
Древнетюркский словарь (1969). Ленинград: Наука.
Жабагин, М.К., Дибирова, Х.Д., Фролова, С.А. и др. (2014). Cвязь изменчивости Y-хромосомы и родовой структуры: генофонд степной аристократии и духовенства казахов. Вестник Московского университета. Серия 23: Антропология. 1. 96–101.
Жабагин, М.К., Сабитов, Ж.М., Агджоян, А.Т. и др. (2016). Генезис крупнейшей родоплеменной группы казахов — аргынов — в контексте популяционной генетики. Вестник Московского университета. Серия 23: Антропология. 4. 59–68.
Жабагин, М.К. (2017). Анализ связи полиморфизма Y-хромосомы и родоплеменной структуры в казахской популяции. Диссертация на соискание ученой степени кандидата биологических наук.
Картаева, Т.Е. (2010). Этногеография казахов нижнего течения реки Сырдарьи (вторая пол. ХIХ–нач. ХХ вв.). Вопросы истории Кыргызстана. 3. 131–139.
Кият, А., Сабитов, Ж.М. (2024). Казахские Шибаниды (XVIII–XIX вв.) в дореволюционных источниках. Былые годы. 19 (4). 1513–1522.
Кузнецов, П. (1900). О таджиках Ташкентского уезда (краткий отчёт). Известия Туркестанского отдела Императорского Русского географического общества. II. 2. 31–51.
Маликов, А.М. (2014). Ходжи Южного Казахстана: история и идентичность. Этнографическое обозрение. 4. 35–45.
Мургабаев, С.С., Малдыбекова, Л.Д., Бахтыбаев, М.М. и др. (2022). История орошения Сыганака. Поволжская Археология. 2 (40). 206–214.
Нагаминэ, Х. (2020). Сыгнак как «порт Дашт-и Кыпчака» и «город-мавзолей»: период от правления левого крыла Джучидов до основания «казахского ханства». Золотоордынское обозрение. 8. 3. 533–551.
Радлов, В.В. (1989). Из Сибири. Страницы дневника. М.: «Наука».
Рожанский, И.Л. (2018). Славянские супер-ветви: Y-ДНК как маркер ранних миграций славян. Исторический формат. 1–2 (13–14). 59–79.
Сабитов, Ж.М., Тажигулова, И.М., Балановский, О.П. и др. (2012). Генеалогия казахстанских Чингизидов (Тука-Тимуридов И Шибанидов) в контексте данных популяционной генетики. Средневековые тюрко-татарские государства. 4. 121–125.
Сабитов, Ж.М., Акчурин, М.М. (2014). Генеалогии (шежире) и генетические данные по происхождению постордынской родоплеменной аристократии. Средневековые тюрко-татарские государства. 6. 127–139.
Сандыбаева, А.Д., Динашева, Л.С. (2025) Население Туркестанского региона в XVIII– начале XX вв.: культура, социальные и политические институты. Известия Национальной Академии наук Кыргызской Республики. 1. S1. 125–131.
Схаляхо, Р.А., Жабагин, М.К., Юсупов, Ю.М. и др. (2016). Генофонд туркмен Каракалпакстана в контексте популяций Центральной Азии (полиморфизм y-хромосомы). Вестник Московского университета. Серия 23: Антропология. 3. 86–96.
Тюрин, А.М. (2024а). Элементы этногенеза хазарейцев по данным популяционной генетики. Nomadic civilization: historical research (Кочевая цивилизация: исторические исследования). 1. 37–49.
Тюрин, А.М. (2024б). Структура субкладов гаплогруппы Y-хромосомы С как свидетельство миграций монголов в XIII, XVII И XVIII вв. Nomadic civilization: historical research (Кочевая цивилизация: исторические исследования). 3. 41–50.
Тюрин, А.М. (2025а) Этническая идентификация казахов по субкладам гаплогруппы Y-хромосомы C2. Nomadic civilization: historical research (Кочевая цивилизация: исторические исследования). 2. 20–39.
Тюрин, А.М. (2025б) Реконструкция элементов этнического прошлого кряшен по естественнонаучным данным. Nomadic civilization: historical research (Кочевая цивилизация: исторические исследования). 1. 9–32.
Турсунов, Б.Р. (2019). Начало покорения Кокандского ханства Российской империей (1852–1863 гг.). Ученые записки Худжандского государственного университета им. академика Б. Гафурова. Серия гуманитарно-общественных наук. 4 (61). 17–23.
Юсупов, Ю.М., Асылгужин, Р.Р., Сабитов, Ж.М. (2023). Башкирские ете ырыу и казахские жетыру по данным этногенетических легенд и преданий и по результатам генетических исследований. Проблемы востоковедения. 4 (102). 22–28.
Balanovsky, O., Zhabagin, M., Agdzhoyan, A. et al. (2015). Deep phylogenetic analysis of haplogroup G1 provides estimates of SNP and STR mutation rates on the human Y-chromosome and reveals migrations of Iranic speakers. PLoS One. 10(4).
Haber, M., Platt, D.E., Ashrafian Bonab, M., et al. (2012). Afghanistan’s Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events. PLoS One. 7(3).
Khussainova, E., Kisselev, I., Iksan, O. et al. (2022). Genetic relationship among the Kazakh people based on Y-STR markers reveals evidence of genetic variation among tribes and zhuz. Frontiers in Genetics. 12.
Liu Shuhu, NIZAM Yilihamu, RABIYAMU Bake et al. (2018). A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP[J]. Acta Anthropologica Sinica. 37(01). 146–156.
Lkhagvasuren, G, Shin, H, Lee, SE et al (2016) Molecular genealogy of a Mongol Queen’s family and her possible kinship with Genghis Khan. PLoS ONE.11. e0161622.
Malyarchuk, B., Derenko, M., Wozniak, M., Grzybowski, T. (2012). Y-chromosome variation in Tajiks and Iranians. Ann. Hum. Biol. 40. 48–54.
Maróti, Z., Neparáczki, E., Schütz, O. et al. (2022). The genetic origin of huns, Avars, and conquering Hungarians. Current Biology. 32. 13. 2858–2870.e7.
Neparáczki, E., Maár, K., Török, T. et al. (2019). Y-chromosome haplogroups from Hun, Avar and conquering Hungarian period nomadic people of the Carpathian basin. Scientific Reports. 9. 1. 165–69.
Wei, LH., Yan, S., Lu, Y. et al. (2018). Whole-sequence analysis indicates that the Y chromosome C2*-Star Cluster traces back to ordinary Mongols, rather than Genghis Khan. Eur J Hum Genet. 26(2). 230–237.
Wen, SQ, Yao, HB, Du, PX et al. (2019). Molecular genealogy of Tusi Lu’s family reveals their paternal relationship with Jochi, Genghis Khan’s eldest son. J Hum Genet. 64(8). 815–820.
Y-DNA Haplogroup C and its Subclades — 2019–2020. https://docs.google.com /spreadsheets /d/ 1XTMjVnybYFfj4mL1UwzDACTy9fZoJdCbENwdfvKWETQ/ edit?gid=928240711#gid= 928240711 (Дата обращения 01.12.2025).
YTree v11.05.00 (29 сентября 2025). https://www.yfull.com/tree/ Дата обращения 01.12.2025 г.
Zerjal, T., Xue, Y.L., Bertorelle, G. et al. (2003). The genetic legacy of the Mongols. Am. J. Hum. Genet. 72. 717–721.
Zhabagin, M., Balanovska, E., Sabitov, Z. et al. (2017). The connection of the genetic, cultural and geographic landscapes of Transoxiana. Sci. Rep. 7 (1). 3085.
Zhabagin, M., Lan-HaiWei, Sabitov, Zh. et al. (2022). Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosome from Central Asia. Genes. 13. 1776.
Zhabagin, M., Bukayev, A., Dyussenova, Zh. et al. (2024). Y-chromosomal insights into the paternal genealogy of the Kerey tribe have called into question their descent from the stepfather of Genghis Khan. PLoS ONE. 19. 9. С. e0309080.
Тюрин, А.М. (2025) Этническая идентификация предков аристократических родов казахов – кожа и торе. Nomadic civilization: historical research (Кочевая цивилизация: исторические исследования). 4. 49–72. https://www.nomadic-kalmsu.ru/jour/article/view/168
Ранее на АШ рассмотрена статья:
Тюрин А.М. Этническая идентификация казахов по субкладам гаплогруппы Y-хромосомы C2. Nomadic civilization: historical research. 2025;5(2):20-39. https://doi.org/10.53315/2782-3377-2025-5-2-20-39 https://www.nomadic-kalmsu.ru/jour
https://aftershock.news/?q=node/1528898
Она опубликована и на сайте «Золотая орда»: https://zolord.ru/articles/3367
Просьба. Подскажите пожалуйста форумы, на которых обсуждаются данные популяционной генетики.
На Еврозийском историческом сервере забанен
На Молекулярной генеалогии у меня персональный пожизненный бан.
Генофонд РФ не работает.
Комментарии
Спасибо за экскурс историю древних казахов!
Что значит это исследование для истории и политики современного Казахстана!? Опять придётся переписывать учебники истории!?
Проблема с торе (они потомки разных родителей) решена легко - адюльтер. В шежере - достоверная информация. Сомнению не подлежит. Хуже с кожа. Мои результаты указывают на массовую фабрикацию генеалогических легенд. Можно предположить, чтоо в это же время осуществлена и массовая фабрикация шежере торе.
Может ли быть так, что, если Ваши результаты конфликтуют со всеми остальными - проблема в Ваших?
У меня нет результатов популяционной генетики. Я интерпретирую результаты тестирований, выполненных специалистами. Результаты моей интерпретации не конфликтуют "со всеми остальными". "Остальные" интерпретировали блоки информации. А я собрал почти всё, что имеется по торе и кожа.
Если у рода Кожа преобладает индоевропейская гаплогруппа R1a1-Z93 (такая же как у скифов, иранских и индийских ариев), то почему так прямо и не написать, а не придумывать им каких-то "таджиков" в предки? (таджики вообще поздний этнос). Вот это, кстати, проблема "популяционной генетики". То, о чем Клесов пишет прямо в своих ДНК-генеалогиях, она исходя из каких-то западных политкорректных соображений всегда пытается сделать максимально мутным, притащить за уши какие-то современные государства и недавно сформировавшиеся нации.
Я строго придерживаюсь тех фактических данных, которые принял во внимание. Кроме того, до R1a1-Z93 гаплогруппа R1a типизирована не во всех выборках.
Недавно опубликован большой массив данных по киргизам. А у них R1a1-Z93 по частотам доминирует. Имею планы эти данные посмотреть.
Совсем, чтоли, писать нечего?
после того как в россии сто лет жгли разрядные книги (начиная с рюриковичей и заканчивая петром первым) официальная история происхождения родов 90% российского мелкого и среднего дворянства 100% миф. а уж генетика... адьюльтер - рулит! читал как-то исследование днк проведенное на нескольких поколениях владимиро-суздальских князей (кажется владимиросуздальские, точно не помню, но какая-то близкая родня мономахам была) - 30% правнуков по прямой МУЖСКОЙ линии никакого отношения к прадеду уже не имела. как так получалось в обществе где "баб взаперти по теремам держали" абсолютно непонятно, но факт. так что пофиг, ни кто учебники переписывать не будет.
По данным популяционной генетики получилось, что еврейки являются потомками женщин всей Западной Евразии а генохронологическое датирование евреев мужчин даёт возраст их общих предков немногом больше 1000 лет. Ничего плохого для евреев не случилось. Никто и ухом не повёл. Лениво попытались сконструировать фальшак - скорость мутаций по Животовскому. За явной дебильности от него отказались.
Масштабные исследования выполнены по генеалогии мак-Дональдов. Кого только среди них не оказалось.
с евреями там вообще прекрасное: 60% ашкеназов произошли от группы женщин-неевреек родом из центральной италии. так что палестинцы (те же друзы, имеющие до 50% сходства своего днк с финистимлянами) имеют прав на земли современного израиля побольше чем собственно евреи
Это никак не повлияло на определение настоящих евреев по матери.
ну, если бы не "пустомудый пророк эзра", тот который и ввел такое лихое определение, евреи до сих пор определяли бы национальность "по папе". но это уже их внутренние разборки.
Кстати, проверка скорости мутаций за последние 100 лет показала, что реальная скорость мутаций примерно в 20 раз выше, чем принято считать. Это означает, что история растянута и она короче примерно в 20 раз. Андрей Степаненко открыл, что до примерно 1838 года детоисчисление вели в месяцах по древнеримской восьмиричной системе. А в современном летоисчислении их интерпретировали как десятичные годы. И поэтому истоия оказалась растянута в среднем в 19.5 раз. Получается Ной жил не 960 лет, а 960 месяцев, т.е. 80 лет. Для тех времен достаточно долго, но для нас нормально. А Сара родила в 96 лет - это 96 месяцев = 8 лет. Время считалось от замужества. Т.е. 8 лет не могла зачать, отчаялась, а потом забеременела. И это уже нормально, в отличие от беременности в 96 лет. Проблема в интерпретации. Столетняя война - 5-8 лет, 300 лет ига - 15-25 лет.
Казахи и пиндосы - братья навек.
... древние укры.
пиндосы не этнос, а результат эксплуатации мирового
отхожего местаплавильного котла -экскрементэксперимент своего рода...Автору это не мешает.
Казахи это северные киргизы, а киргизы это южные казахи.
КАзахи в основной своей массе говорят на кыпчакских диалектах, а кыргызы используют южно-сибирские диалекты и родственны хакасам (хагяс-кыргыз)
Ну и за что любить А. Клёсова казахам? Не ту песню от него ждали.
А Клёсов сделал много полезного. Но он не имеет ни малейшего отношения к данным, приведённым в моей статье. Кстати, про него в последнее время ничего не слышу.
Публикации 2020-2025 годов - https://dna-academy.ru/papers/
ДНК замечательных людей - https://topdnk.ru/
Значит ошибся про А.Клёсова. Бывает. Может поэтому и не слышите, что его исследования не подтверждают мифов народов о себе славных. Вот если-бы его Академия направо-налево раздавала документы, красивые да с печатями, подтверждающие самописные учебники истории величайших народов. Тогда да, был бы на слуху.
Угу. И попгенетикам тоже...
Попгенетики из кое-чего высосут что угодно.
«История скифов» глазами «геномных попгенетиков»
А. Клёсов не владение технологией работы с полногеномными аутосомными маркерами. А это мощнейший инструмент популяционной генетики.
Дичь какая, такое пристальное внимание к родам, Чингисханам, Мухамедам и ДНК, маркер отсталой/мертвой банановой республики. В живом обществе люди считают своими тех у кого в голове те же ценности и основы, и неважно он нерг, якут или русский.
Нужно отвыкать оценивать другие сообщества по своим критериям.
Поясните пожалуйста ещё вот что. Эти княжеские роды ведут свою родословную и от Чингисхана, и от Мухаммеда. Но последний сказал "Если вы услышите, как кто-то возносится своим происхождением, как это было во времена джахилии, то скажите ему, чтобы он схватил за член своего отца. И не говорите это другими словами!" Причём последнее предложение явно предписывает даже не пытаться передавать этот (достоверный) хадис иными словами. Поэтому и привожу цитату так грубо, как в оригинале. Почему они не пытаются стереть свою историю и культуру в этой части, как предписывает их религия? У них своя версия её? Или это такая форма лицемерия?
Вы конечно правы, что для работы с другими сообществами "через пряник", это строгонеобходимо, но есть ли успешные примеры работы с банановыми через пряник? А для работы через кнут, плевать на все тонкости.
С чего вы решили, что народы с пережитками родо-племенного строя мыслят как современные люди? Не знаю лучше ли общество современно мыслящих "снежинок" или либерастов, но родо-племенное тоже имеет право на существование.
Вопросы крови - самые сложные вопросы в мире!
....Как причудливо тасуется колода! (с)
Да. Для меня это интересное занятие - вычисление кто и как оттусовался.
Судя по истории вопроса наследники Тимучина уже какбэ не родные, а приёмные. А кто там папа генетический поди узнай. И непонятно, учитывая это, есть ли смысл так дотошно выискивать генетику у Торе? А из бесед, шежире (генеалогическая родословная) казахов лично для меня тёмный лес. Но они сами как-то там ориентируются. Причём ориентируются уверенно и самое удивительно что практически все. Спроси какой у вас РУ, ответ будет исчерпывающим и подробным. Такая вот устная историческая память. Впрочем не историк и не этнограф, просто была возможность интересоваться вопросом, география командировок позволяла.
"Кожа, в которой я живу", 2011.
Учебники истории переписывать не стоит. Просто нужно разрешить включать в них попаданцев и публиковать как произведения интеллектуально-развлекательной литературы. Предмет история после этого изучать в процессе изучения литературы
Бывшая партноменклатура начала создавать себе вековую родословную от самых истоков.
В январе 2022 года уже была волна по поводу жузов, но там нет славных предков, а только общины.
ИзТория - оружие по высшему приоритету воздействия на общество.
Махинации с изТорией, псевдо-научные обоснования - это часть "гибридной" войны!
Посмотрите на хохлостан! За 30 лет вырастили манкуртов. Казахи дружным строем шагают по этому же пути!
Это то понятно. Великая степь она же и великая дорога переселения народов, место кочевого скотоводства, где перемешивание неизбежно. Это как у х0хлов, там же тоже западная часть той же степи...
Теперь осталось провести исследование на тему "аристократов" из Папуа -Новой Гвинеи или ещё каких нибудь людоедов.
Это важно.
По преданиям, исторический центр формирования народов мандинка находился в верховьях реки Нигер, где в VIII веке сложилось политическое объединение мандинка — империя Мали. Сами мандинка считают себя потомками легендарного основателя этого царства.
Палат 6 очень много на Планете
Так нет проблем. Правительство Папуа-Новой Гвинеи может заказать такое исследование генетикам.
То есть за это ещё и заплатили
Чудный, чудный мир
Ну да. Генетические исследования казахов выполняются, в основном, за деньги Казахстане.
Хорошо, когда денег много в стране
Как понимаю, результаты исследований они же заказывают. Какой смысл вообще что-то городить, если беспристрастности в этом вопросе не было никогда и ни у кого?
Вся эта генетика родов и галогрупп - сплошная политика и словоблудие. Нарисуют любых арийцев на своей мульке, может марсиан найдут или богов каких-нить....
Возможно, что для этого вся эта вахканалия и проводится. Чтобы окончательно дискредитировать эти родоплеменные. Жаль, что медициа при этом страдает, РНК-вакцины и пр. передок науки.
Вы путаете генетиков мс историками. Генетики - это биологи. У них всё честно. Генетики друг за другом приглядывают.
Серер — потомки легендарного народа гангара. Под натиском Альморавидов переселились к югу от реки Сенегал и совместно с фульбе в 1186 г. основали государство Вало. Впоследствии переселились в низовья Сенегала и независимо от фульбе основали самостоятельные государства Син и Салум.
Страницы