






В первой из двух предыдущих статей я попытался описать сильно упрощенный набор терминов и понятий из общей теории систем (ОТС), которые применил во второй статье серии для поиска подобия социальных систем мира людей и мира животных максимально низкого ранга.
Гипотеза, которую я описываю в серии, состоит в том, что согласно ОТС одноранговые системы разных иерархий систем (живых, искусственных, социальных, химических и т. д.) ведут себя подобным образом. Найдя некоторое соответствие в рангах двух систем (биологической и социальной), и взяв их за отправную точку, я попытаюсь выявить общие характеристики развития систем.
Цель достаточно понятная - описать то, что происходит сейчас в мире людей с точки зрения законов развития систем, чем это грозит человечеству, хотя бы в общих чертах.
В предшествующей статье я описал часть совпадений в системных свойствах двух систем: биологической системе колониального типа одноклеточных зеленых организмов вольвокс и социальной системе первобытных родовых обществ. Это позволяет предположить их одноранговость.
Динамическая устойчивость этих систем не вызывает сомнений, т. к. вольвокс достаточно распространенный род колоний в стоячих пресноводных водоемах, а родовые общины по имеющимся данным палеонтологов были широко распространены в прошлом и численность людей к расцвету данного типа социальных систем достигала приблизительно 5 млн. человек.
Однако, не ясно была ли данная динамическая устойчивость получена сразу или это результат определенных изменений в системах.
Ответ на этот вопрос сравнительно недавно был найден и продемонстрирован в результатах 5-ти летнего эксперимента, проведенного группой ученых из новозеландского института передовых технологий, немецкого института эволюционной биологии Макса Планка и ученых из США. Результаты были опубликованы в журнале Nature №7525 в 2014 г. в статье «Life cycles, fitness decopling and the evolution of multicellularity».
В эксперименте исследовались колонии бактерий Pseudomonads fluorescens, которые тоже, как и вольвокс, жгутиковые, и при определенных типичных мутациях могут выделять клей. Обычно они одиночки, но клей позволяет им образовывать колонии в виде матов (пленок в несколько слоев) на поверхности воды. При этом верхний слой получает доступ к атмосферному кислороду, который обычно им не доступен. Это образует в сочетании с пищей, распределенной в воде, условия для процветания колоний и быстрого размножения, как составляющих их бактерий, так и колоний.
При этом в них формируется специализация бактерий. Одни поставляют кислород, другие добывают и транспортируют пищу, третьи распределяют потоки. Исследователи обнаружили, что во внутренних слоях матов в части колоний существуют бактерии, которые не выделяют клей, и фактически паразитируют на других. Все дело в том, что производство клея для этих бактерий достаточно энергозатратная процедура. Минимизируя свои затраты бактерии-паразиты (правда исследователи их мягко назвали жуликами) размножаются гораздо быстрее своих собратьев. В результате маты разрушаются и тонут. Но мало того, бактерии, образовывавшие их гибнут тоже, в т.ч. и паразиты, т. к. не способны вновь адаптироваться к одиночному существованию.
Кроме колоний, в которых паразиты имели возможность беспрепятственно размножаться, были колонии, в которых таких бактерий не было в принципе или бактерии-паразиты были как-то адаптированы к нуждам колонии.
На ряду с тем, что они оказались динамически устойчивыми, у них были и существенные отличия. Колонии с адаптированными бактериями-паразитами в целом быстрее адаптировались у изменению условий внешней среды, но при разрушении происходила гибель всех организмов.
Колонии с отсутствием бактерий-паразитов медленнее адаптировались к изменениям внешней среды, но при разрушении колоний демонстрировали высокую одиночную выживаемость, т. е. их индивидуальная адаптивность сохранялась в условиях колоний.
У этого явления есть вполне рациональное объяснение. В 80-х годах прошлого века для растительных и живых организмов был сформулирован принцип энергетического минимума в индивидуальном развитии, согласно которому минимальные энергозатраты наблюдаются в зоне наиболее благоприятных для развития организма значений факторов среды и соответствуют наиболее устойчивому состоянию организма (гомеостазу).
Принцип индивидуальной минимизации затрат является общим для всех живых организмов. В силу этого принципа все, что не используется, отмирает. Так отмирают неиспользуемые органы, например, хвост у человека или незадействованные долгое время в передаче сигналов синаптические связи, из-за чего мы многое забываем. В мозгу человека есть целый сложный механизм удаления таких связей.
Этот принцип является эквивалентом принципа минимума общей потенциальной энергии связей частиц в физических и химических системах.
С точки зрения ОТС этот принцип означает минимизацию структуры потоков. Однако, это не возможно без изменения структуры элементов. А элементы — это определенные функции системы, и упрощение структуры — это исключение функций, что для одиночных организмов означает смерть.
Для бактерии во внутренней части колонии эта колония является окружающей средой с максимально благоприятными условиями существования. Таким образом, в условиях халявных потоков происходит естественным образом минимизация функций в общем-то обычных бактерий, не наделенных даже в принципе толикой разума. В силу того, что это принцип индивидуального применения для систем, то бактерия реализует ее в версии уменьшения выходных потоков энергии, вещества, информации. При этом каждая бактерия-паразит генерит энтропию в связанные бактерии, находящиеся с нею на связи, т. к. перекрывает им жизненно важные потоки. Это тоже влияет на структуру этих бактерий, т. к. в них разрушаются элементы, работавшие на этих потоках.
Высвобожденные потоки бактерии-паразиты усиленно расходуются на размножение. В какой-то мере люди для смежных систем являются паразитами, которые до недавнего времени бурно размножались.
Экспериментаторы использовали селекционные методы для ускорения эволюционных процессов, и в результате получили еще один интересный вывод. Колонии, зараженные внутренними бактериями-паразитами, превратили их в эффективный аппарат размножения колоний. Эти бактерии образовывали миниколонии, которые затем отделялись от родительской колонии и начинали самостоятельное развитие. Условия в миниколонии далеки от максимально благоприятных условий сложившейся колонии и соответственно не происходит упрощения структур. Такой же механизм применяли и продолжают применять живые и социальные системы колониального типа. По-видимому, это было проявлением механизма формирования системы дифференцировки клеток в его первоначальном варианте.
В силу интересных особенностей языка название колониальная система получила и система внешнего паразитирования, когда есть внешний по отношению ко всем элементам системы паразит (метрополия), которые питается потоками всей колониальной системы.
В биологии паразит так и определяется, как нечто, что питается соками тела организма, тканями или переваренной пищей хозяина-носителя.
Почему я употребил понятие «нечто» в отношении паразитов. Все дело в том, что существует класс паразитов, которые не относятся к живым организмам. Это вирусы. Их особенность в том, что в отличии от живых систем, которым для существования, т. е. динамической устойчивости и сохранения целостности, требуется постоянный и значительный поток энергии, у вирусов существует режим статической устойчивости. Он позволяет им достаточно долго избегать разрушения между состояниями динамической устойчивости, когда он попадает в организм-хозяина и питается его внутренними потоками.
Это объяснимо тем, что функций кроме размножения у него нет, соответственно механизм работает только на эффективное размножение в хозяине, т. е. минимален по затратам энергии. Основная биологическая функция при это выполняется лучше некуда.
Существуют паразиты и в технических системах. К ним относят механизмы, которые приводят к потерям потоков энергии, вещества, информации (энтропии). Например, дефекты в трубопроводах или проводниках, приводящие к утечкам и пробоям, или механизмы коррозии, утечек магнитных полей, тепла в зданиях и т.п.
Скорость размножения паразитической структуры зависит от величины замкнутых на нее потоков. Быстрое размножение — нарастание замкнутых на паразита потоков. При значительном превышении скорости роста паразита по сравнению с системой-хозяином наступает момент, когда отбор потоков превышает порог динамической устойчивости системы-хозяина, и она разрушается.
Поэтому одной из существенных составляющих стратегии выживания многих паразитов является стратегия повышения скорости размножения систем-хозяев и повышения эффективности работы каналов преобразования потоков. Известно, что существует обратная зависимость от распространенности паразитов и уровней заболеваний астмой и аллергиями. На островных поселениях выявлена прямая зависимость фертильности женщин от зараженности популяции отдельными видами паразитов.
Вот здесь и возникает точка, где паразиты могут превратиться в симбиотов. Как правило, все нонешние симбиоты человека являются бывшими паразитами. Так геном человека на 80% состоит из отрезков геномов вирусов и других паразитов. Остальные считаются принадлежащими человеку только на том основании, что они ответственны за дифференцировку клеток, из которых формируются наши органы. Однако, сам переход к дифференцировке в многоклеточных организмах по моему личному мнению является следствием непрерывной борьбы организмов с паразитами. Например, те же гены, которые запускаются (экспрессируются) в клетках для получения нужной для конкретного органа конфигурации, перемешаны в геноме. Но существуют гены-инспекторы, которые достались нам от паразитов, которые контролируют этот процесс и блокируют не имеющие значения паразитические цепочки.
Особенно важны результаты, ведущие к повышению эффективности преобразования потоков в системе, т. е. повышению ее коэффициента полезного действия. С точки зрения развития это выгодно с двух взаимосвязанных сторон. Улучшение ведет к повышению эффективности, повышение эффективности ведет к получению энергии для совершенствования системы в условиях достаточно постоянной или падающей мощности потоков, что происходит гораздо чаще.
Существует еще аспект развития, который играет ключевую роль в иерархии систем. Изменения в организмах накапливаются постоянно под действием колебаний потоков, поступающих из внешней среды, колебаний балансов входных и выходных потоков, влияния паразитов. Однако, закрепление изменений на уровне родовых признаков системы происходит только во времена точек бифуркации, когда одномоментно отсекаются все системы, не имеющие изменений, необходимых для выживания в новых условиях. Напомню про «бутылочные горлышки» в развитии человеческих предков, о которых я писал в предыдущей статье серии.
Кроме того, существует парадокс принципа минимизации затрат, который почему-то в случае паразитов ведет к упрощению, а в случае их жертв к усложнению. Это возможно, если действие паразита длится меньше, чем период, необходимый для разрушения системы. Такое действие является аналогом изменений балансов потоков, и работает как стимул к повышению эффективности путем усложнения. За примером ходить далеко не надо. Выработка иммунитета к вирусным и бактериальным заболеваниям при вакцинации является именно таким результатом.
С точки зрения ОТС паразит — это некая система ранга элементов другой системы, которая встраивается в структуру потоков энергии, вещества, информации (энтропии) между элементами этой системы и отбирает часть этих потоков, ухудшая баланс потоков в системе. Если механизмы поддержания динамической устойчивости справляются с проблемой, то система сохраняет целостность, в противном случае разрушается.
Стратегий борьбы не очень много. Уничтожить, приспособить паразита или смешанный. Причем речь не идет о разумности действий этого механизма. Все должно выполняться автоматически или под действием инстинктов в лучшем случае.
В частности можно предположить, что в колониях вольвоксов обычная клетка, попадая в условия максимального потока, превращалась в паразита. Соседние клетки прекращали связи с нею. В результате для сохранения входящего потока клетка перемещалась к следующим «жертвам». Там происходило то же самое. И так до тех пор, пока клетки-паразиты не собирались в миниколонию, которую родительская колония благополучно выкидывала наружу. А в условиях внешней среды уже паразитировать не приходится.
Практически тоже самое происходило в родовых обществах. Особо умные и наглые, для кого табу не табу, получали путевку в жизнь. А поскольку они, как правило, имели сторонников, то эти группы и становились новыми родовыми обществами, которые вынуждены были уже сами существовать по знакомым правилам и табу, чтобы выжить.
Подход совмещает и стратегию уничтожения при неудаче новой колонии и стратегию приспособления в виде системы размножения колоний при удаче.
Возвращаясь к теме развития социальных обществ необходимо отметить, что гипотеза возникновения механизма дифференцировки находит хорошее подтверждение в истории человечества.
Родовые общины охотников и собирателей быстро размножаясь и распространяясь по наиболее благоприятным зонам берегов рек и морей, прежде всего, субтропических регионов очень быстро стали подрывать кормовую базу. Имеющиеся к тому времени технологии земледелия позволяли решить эту проблему, хотя бы в основном. Но это требовало оседлого проживания и изменения условий существования системы.
Закрепление на территориях приводило к непрерывному увеличению стычек между родовыми группами и их племенными объединениями. В результате были победители и побежденные. Их объединения превращались в более сложные и высокоранговые социальной системы — государства.
Существование государств не возможно без управления, силовых структур, городов-поселений, официальных религий, законов и их толкователей и других органов социальной системы. Кроме этого, возникло разделение на классы, поскольку потоки пошли через отдельные социальные группы победителей и автоматически обычные люди из бывших родовых общин победителей перешли в режим паразитарного существования. Для этого им даже думать не пришлось. Победил и греби к себе, ты ж не курица.
Именно для существования этих групп нужны были специализированные органы социального устройства. Для занятия места в этих органах создавалась и реализовывалась все более сложная система дифференцировки людей по признакам принадлежности к определенным слоям и по подготовке к работе в этих органах. Солдата, писца, церковного служащего, судью и т. д. надо учить, назначать, обеспечивать соответствующими атрибутами и выстраивать отношения с другими группами и людьми.
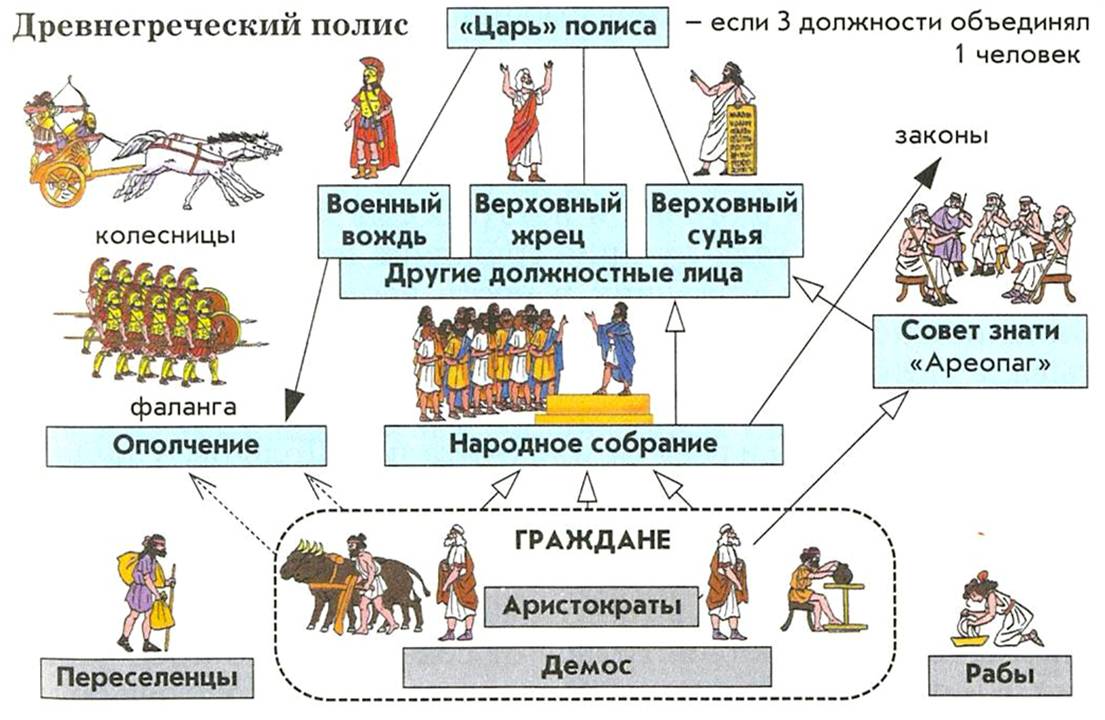
Это и есть дифференцировка людей в социумах, являющаяся аналогом дифференцировки клеток в сложных многоклеточных организмах.
Древние государства стали первыми сложными социальными системами с дифференцировкой элементов (органов) по функциям, со своим сложным внутренним устройством, со своими тараканами, т. е. паразитами, как внешними, так и внутренними.
Комментарии
Спасибо!
На здоровье.
продолжаем наблюдение.
просто вроде текст (вашей имхи) есть, а комментить нечего. нету тезисов для зацепки..