






Много людей использует термин ДНК. Но статей, нормально описывающих, как она работает почти нет (понятных не биологам). Я уже описывал в общих чертах устройство клетки и самые основы ее энергетических процессов. Теперь перейдем к ДНК.
ДНК хранит информацию. Это знают все. Но вот как она это делает?
Начнем с того, где она в клетке хранится. Примерно 98% хранится в ядре. Остальное в митохондриях и хлоропластах (в этих ребятах протекает фотосинтез). ДНК — это огромный полимер, состоящий из мономерных звеньев. Выглядит примерно так.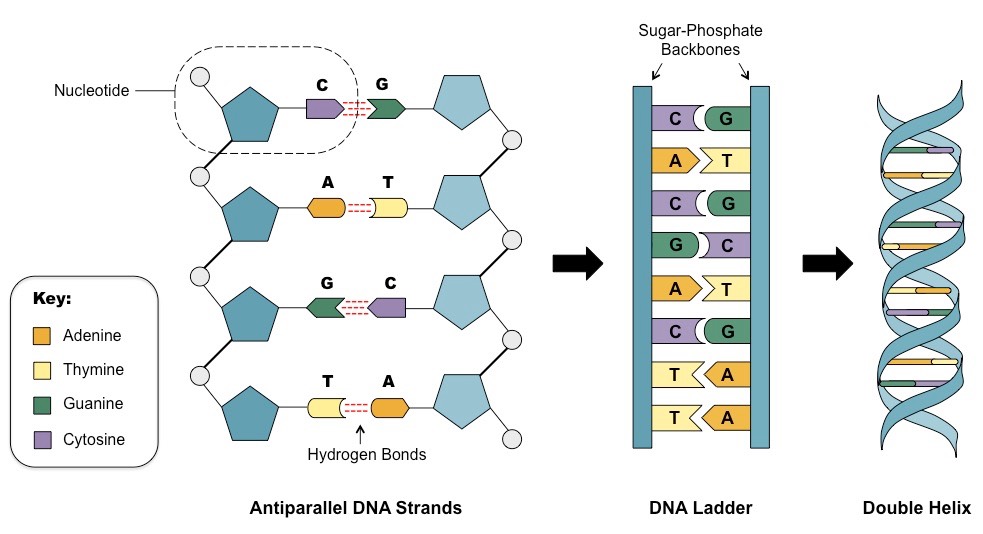
Что мы тут видим? Во-первых ДНК — двухцепочечная молекула. Почему это так важно — чуть позже. Далее мы видим синие пятиугольники. Это молекулы дезоксирибозы (такой сахар, чуть меньше глюкозы. От рибозы отличается отсутствием одной OH группы, что придает стабильности молекуле ДНК, в отличие от РНК, в которой используется рибоза. Дальше, для простоты опущу приставку дезокси и буду просто говорить рибоза, да простят нас щепетильные товарищи). Маленькие кружкИ — остатки фосфорной кислоты. Ну и собственно есть азотистые основания. Всего их 5, но в ДНК в основном встречаются 4. Это Аденин, Гуанин, Тимин и Цитозин. То есть, есть рибоза с которой связано азотистое основание. Вместе они образуют так называемые нуклеозиды, которые связываются друг с другом с помощью остатков фосфорной кислоты. Таким образом мы получаем длинную цепь, состоящую из мономеров. Теперь посмотрите на увеличенную левую цепь. Видите C и G соединены тремя пунктирными линиями, а T и A двумя. Что это значит? Да, ДНК состоит из двух цепей, но что удерживает их вместе? Есть такая штука, как водородная связь. Выглядит примерно так. На атомы кислорода (O) и азота (N) формируется частичный отрицательный заряд, а на водороде (H) — положительный. Это приводит к формированию слабых связей.
Связи действительно очень слабые. Их энергия может быть в 200 раз ниже энергии ковалентных связей (образуются за счет перекрытия пары электронных облаков, например связь в молекуле CO2). Однако таких связей много. В каждой нашей клетке ДНК цепи связаны почти 16 миллиардами слабых связей, не мало, согласны?
Но вернемся к числу связей между основаниями. Цитозин и Гуанин связаны тремя связями, а Аденин и Тимин — двумя. Это приводит к тому, что Г и Ц связанны куда прочнее, чем А и Т. Некоторым организмам нужна особая стабильность связей ДНК, например живущим при высоких температурах. При нагревании ДНК содержащая больше ГЦ пар более стабильна. Так что хочешь жить в гейзере — имей много ГЦ пар. Хотя последние исследования говорят, что явной связи между GC составом (% ГЦ пар от всех пар) и температурой обитания нет. Стоит сказать, что варьирует он сильно. Так у Candidatus Carsonella ruddii PV (внутриклеточный эндосимбионт) он примерно 16%, у нас с вами почти 41%, а у Anaeromyxobacter K (бактерия вполне себе средних размеров) достигает 75%.
Тут вы можете видеть связь GC состава с размером генома бактерий. Mb — миллион пар нуклеотидов. Показатель довольно вариативный. Его, кстати, часто юзают как фичу при обучении различного рода классификаторов. Сам недавно писал классификатор для распознания патогенов на основе сырых данных секвенирования и оказалось, что GC состав даже по одному риду вполне себе можно использовать.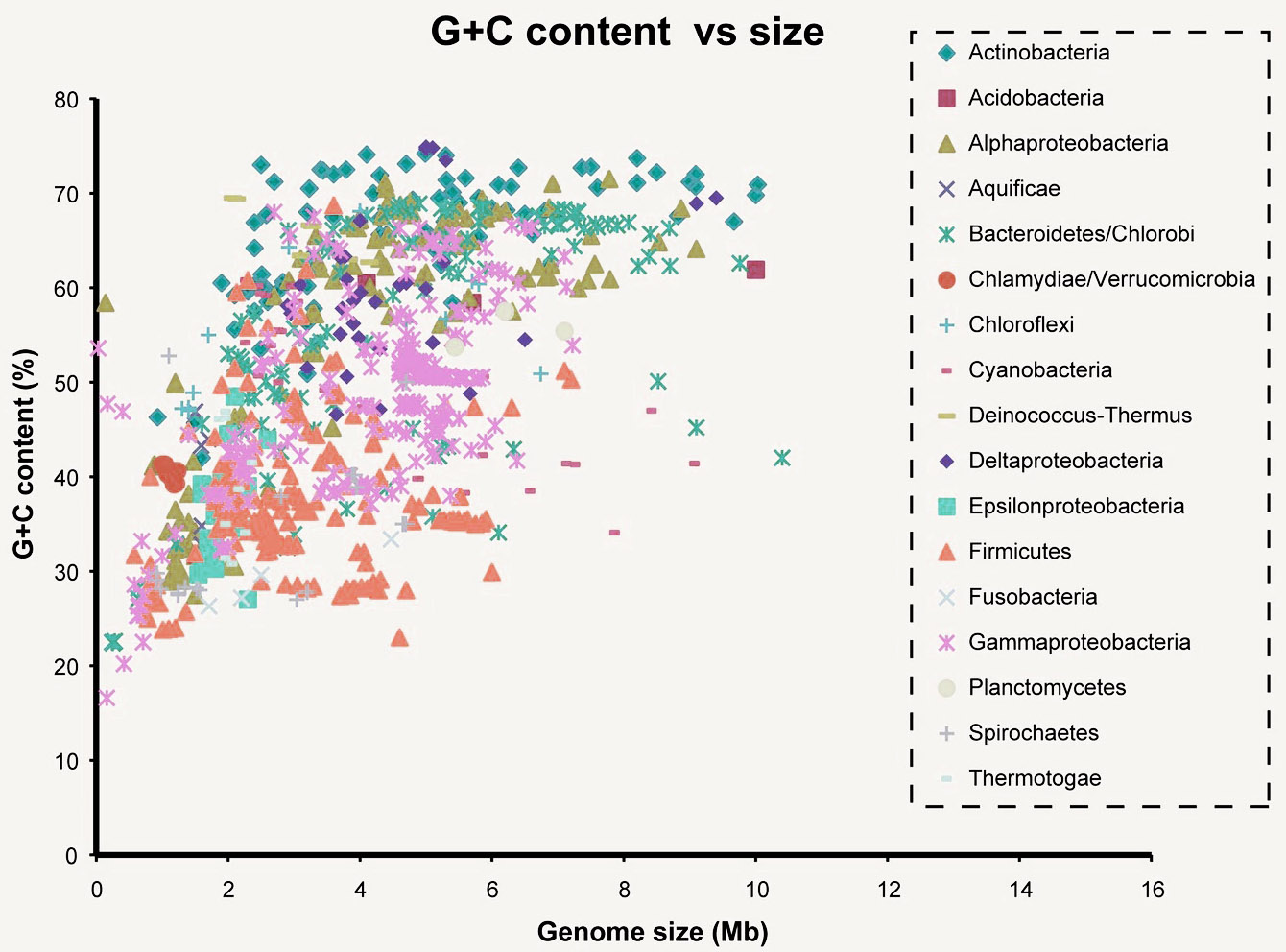
Пока не забыл. Почему важно, что ДНК двухцепочечная? На основе одной цепи можно восстановить другую. Если в одной цепи поврежден кусок напротив последовательности Аденин-Аденин-Цитозин, то мы точно знаем, что до повреждения там был Тимин-Тимин-Гуанин. Таким образом наличие второй цепи позволяет надежней хранить информацию.
Круто! Теперь вернемся к самой молекуле ДНК. Это цепочка из 4х типов звеньев. Однако насколько длинная? У Candidatus Carsonella ruddii PV уже упомянутого выше всего 160 000 нуклеотидов. У нас с вами 3.2 миллиарда (в гаплоидной клетке, то есть с одним набором хромосом. У большинства наших клеток их два). Кажется много, да? На самом деле нет. У одноклеточной амебы (Amoeba dubia) он примерно 670 миллиардов пар нуклеотидов. Кажется что это бесконечно длинная цепочка, поэтому давайте переведем размер в любимые нам метры. Если все наши хромосомы (их 46, не забываем; 23 по две копии на каждую) развернуть и вытянуть в одну линию, получится примерно 2х метровая цепочка. ДНК одной амебы хватит, чтоб опоясать футбольный стадион. Но к чему я веду? Ядро, в котором ДНК хранится не очень большое. У нас оно в среднем диаметром в 6 мкм. Не очень то много, если хочешь свернуть 2х метровую нить, пусть и очень тонкую. Причем нужно не просто запихать нить в ядро. Нужно свернуть таким образом, чтобы в любой момент можно было обеспечить доступ к любому ее участку. Задача сложная. И с ней успешно справляются специализированные белки. Они создают ряд спиралей и петель, которые обеспечивают все более и более высокие уровни упаковки и не до допускают спутывания ДНК в гордиев узел. Давайте поговорим о том, как она упаковывается.
Сразу скажу, упаковывается она очень по разному. Но если откинуть экзотику, то остается два способа. Первый характерен для бактерий, второй для эукариот (или иначе ядерных).
Упаковка ДНК у бактерий
Начнем с братьев наших меньших. Бактерии сами по себе обладают не очень большим геномом, в среднем от 1 до 5 миллионов пар нуклеотидов. Наиболее характерное их отличия от нас в том, что у них нет ядра и ДНК плавает в клетке. Не совсем плавает, оно частично прикреплено к клеточной мембране и тоже свернуто, но не так сильно как у нас.
Второе. Бактериальная ДНК чаще всего кольцевая. Так ее проще копировать (нет концов, которые могут потеряться при копировании и не нужно придумывать механизмы сохранения концов). Обычно такое кольцо одно, но у некоторых бактерий их может быть 2 или 3. Есть еще кольца поменьше (от пары тысяч до пары сотен тысяч остатков).Имя им плазмиды, и это вообще отдельная история.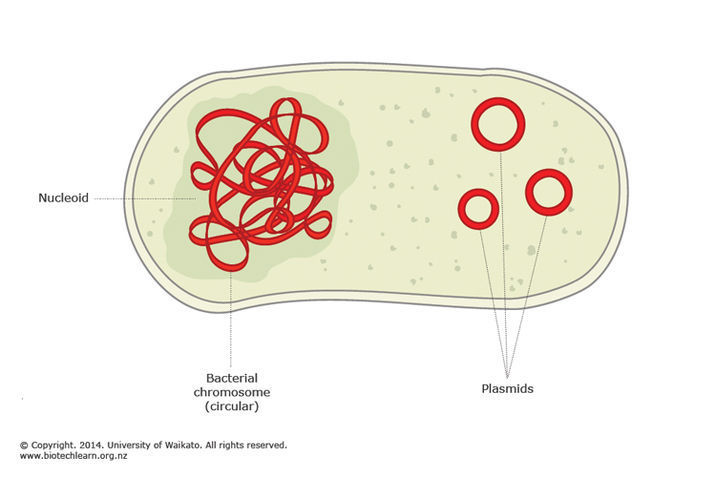
Вернемся к упаковке ДНК. ДНК упаковывают белки-гистоны (есть еще гистоноподобные белки). ДНК это дезоксирибонуклеиновая кислота. Кислота. Это значит что она отрицательно заряжена (за счет остатков фосфорной кислоты). Поэтому белки, связывающие ее положительно заряжены. Таким образом они могут связываются с ДНК. ДНК бактерий вместе с белками ее упаковывающими формируют нуклеоид, при этом на долю ДНК приходится 80% от его массы. Выглядит это примерно так. То есть кольцевая ДНК делится на домены по 40 тысяч пар нуклеотидов. Затем происходит скручивание. Внутри доменов тоже происходит скручивания, но его степень в разных доменах отличается. В среднем степень упаковки бактериальной ДНК варьирует от сотни до тысячи раз.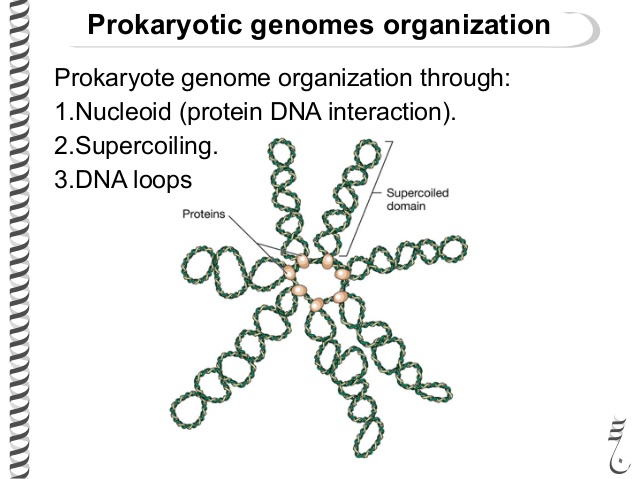
Есть еще прикольное видео.
Упаковка ДНК у эукариот
Тут все куда интересней. Наше ДНК хорошо упакована и спрятана внутри ядра. И она куда эффективней упакована, нежели у бактерий. Во время митоза (деление клетки) размер 22й хромосомы составляет 2 мкм. Если ее распутать и вытянуть, она будет уже 1,5 см. Что соответствует степени упаковки в 10 000 раз. Это около максимальная степень упаковки нашей ДНК. Во время деления нужно максимально упаковать ДНК, что бы эффективно разделить ее между дочерними клетками. В обыденной жизни степень компактизации составляет примерно 500 раз. Со слишком упакованной ДНК сложно считывать информацию.
Есть несколько уровней упаковки ДНК эукариот
Первый — нуклеосомный уровень. 8 белков-гистонов формируют частицу на которую наматывается ДНК. Затем еще один белок ее фиксирует. Выглядит примерно так.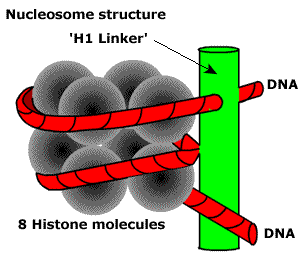

Получаются своего рода бусы. Плотность упаковки благодаря этому возрастает в 7-10 раз. Далее нуклеосомы упаковываются в фибрилы. Немного похоже на солениод. Тут суммарная степень упаковки может достигать 60 раз.
Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. Фибрила разбита на участки по 10 — 80 тысяч пар азотистых оснований. В местах разбивки находятся глобулы негистоновых белков. ДНК — связывающие белки узнают глобулы негистоновых белков и сближают их. Образуется устье петли. Средняя длина петли включает примерно 50 тысяч оснований. Эту структуру называют интерфазной хромонемой. И именно в ней наше ДНК находится большую часть времени. Уровень упаковки здесь достигает 500-1500 раз.
При необходимости клетка может еще больше компактизировать генетический материал. Идет образование более крупных петель из хромомерной фибриллы. Эти петли в свою очередь образуют новые петли (петли в петли… и это не вязание). Которые в конечном счете формируют хромосому.
В целом процесс упаковки можно описать примерно так.
В итоге из нитей ДНК мы получаем, при делении, суперскрученные структуры, которые можно увидеть под микроскопом. Их мы и зовем хромосомами.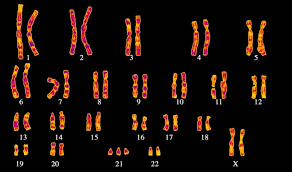
Собственно вещество хромосом зовется хроматином. И степень его упаковки отличается в зависимости от участка хромосомы. Есть эухроматин и гетерохроматин. Эухроматин это довольно расплетенная область хроматина, в ней ДНК находится на хромомерном уровне (упаковка в 500 — 1000 раз). Здесь происходит активное считывание информации. Например, если сейчас клетка активно синтезирует белок А, то область ДНК, его кодирующая будет в состоянии эухроматина, что бы ферменты, «читающие» ДНК могли до нее добраться. Гетерохроматин же содержит ту часть ДНК, которая клетке не особо нужна сейчас. То есть ДНК максимально плотно упакована, дабы не путаться под ногами. В зависимости от потребностей клетки одни области хроматина могут частично расплетаться, в то время как другие — сплетаться. Таким образом еще и осуществляется регуляция (очень грубое приближение), ведь к скрученной области не добраться, и значит ее не прочитать.
Продолжение следует...
Комментарии
Замечательная статья. Интересно.